 药智官方微信
药智官方微信
 药智医械公众号
药智官方微信
药智医械公众号
药智医械公众号
药智官方微信
药智医械公众号
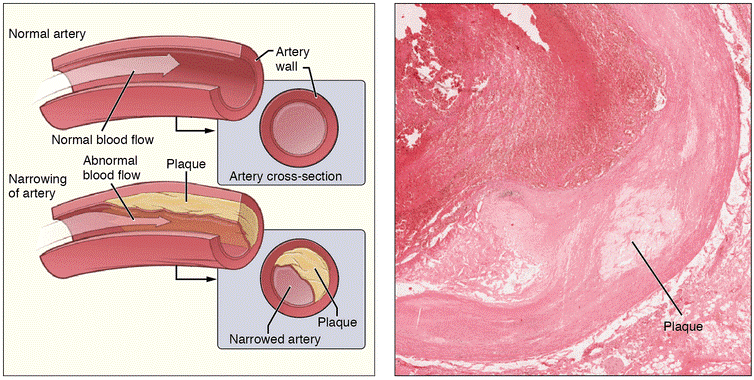
导读动脉粥样硬化(atherosclerosis,AS)是一种缓慢型临床疾病,是心血管疾病的核心,是引起心肌梗死、中风和周围血管疾病的导火索[1]。脂质代谢异常造成的脂质沉积是动脉粥样硬化最直接的致病因素,受累动脉病变从内膜开始,积聚的脂质外观呈黄色粥样,一般先有脂质和复合糖类的异常沉积,进而纤维组织增生及钙质沉着,并伴随着动脉中层逐渐蜕变和钙化,导致动脉壁增厚变硬、失去弹性以及血管腔狭窄。随着血管逐渐狭窄,病变又会累及不同器官,出现头晕、心痛、胸闷和顽固性高血压、糖尿病等症状,可以说动脉粥样硬化就像一颗“不定时炸弹”。目前我国是全球动脉粥样硬化疾病高发区,60岁以上老年人的发病率高达79.9%;并且近几年偏于年轻化趋势,严重影响人类的生命健康。动脉粥样硬化及动脉斑块简图[2]动脉粥样硬化是多因素共同作用引起的,发病机制较为复杂且尚未完全阐明。目前来看,大小鼠是动脉粥样硬化研究中最常用的物种,尽管有局限性,但其与人类的动脉粥样硬化过程的许多关键特征是相似的,仍然是动脉粥样硬化模型的首选物种。动脉粥样硬化的发展与胆固醇运输颗粒低密度脂蛋白(LDL)密切相关[3],主要为载脂蛋白E(ApoE)基因和低密度脂蛋白受体(LDLR)基因,可为动脉粥样硬化的治疗和预防提供参考依据。B-Apoe KO小鼠载脂蛋白E(Apolipoprotein E, ApoE)是一种与脂质颗粒相关的载体蛋白,主要由肝脏和外周组织中的巨噬细胞产生,是血浆脂蛋白的核心成分。ApoE将脂蛋白、脂溶性维生素和胆固醇输送到淋巴系统,然后进入血液,防止富含胆固醇的颗粒在血浆中积聚,是预防、治疗动脉粥样硬化血管疾病的主要靶点基因。研究表明,ApoE能够通过多种方式预防动脉粥样硬化[4]:促进循环系统中富含甘油三酯的脂蛋白的有效吸收;维持正常的巨噬细胞脂质稳态;调节免疫功能等。百奥动物BioMice利用基因编辑技术构建了B-Apoe KO小鼠。利用高脂饮食喂养诱导的小鼠胆固醇水平升高程度加大,硬块的形成进一步加剧,类似于人类动脉粥样硬化及动脉粥样硬化性心脏病中的饮食依赖性,可用于心血管疾病及阿尔兹海默症的研究。B-Apoe KO小鼠特殊饮食诱导动脉粥样硬化模型的建立(A-B)小鼠在12周和20周时的动脉粥样硬化斑块。(C-D)主动脉窦在第12周和第20周的油红结果。(E)12周时主动脉弓大体染色。12周时动脉粥样硬化小鼠模型的组织染色和血液生化结果(A)主动脉窦的H&E结果增加。(B-C)免疫组化显示α-SMA和F4/80的表达。(D)血清ALT、AST、TC、TG、HDL-C和LDL-C水平。数值以平均值±SEM表示。**p<0.01,***p<0.001。B-Ldlr KO大鼠低密度脂蛋白受体(Low density lipoprotein receptor, LDLR)是一种存在于网格蛋白包被的凹坑中的细胞表面受体,介导血浆中主要携带胆固醇的脂蛋白LDL的内吞作用,并且它能够结合APOE蛋白,从而清除血液中的脂蛋白颗粒,调节血浆胆固醇水平[5]。百奥动物BioMice利用基因编辑技术构建了B-Ldlr KO大鼠模型。尤其是当喂食高脂饮食时,这使Ldlr敲除大鼠成为研究动脉粥样硬化的良好模型进而具有重要的临床意义。这种大鼠可用于支持许多领域的研究,包括糖尿病、肥胖研究、过早患冠心病的风险及代谢研究等。B-Ldlr KO大鼠的模型验证及分析纯合Ldlrtm1/Bcgen大鼠在4个月大时表现出显著升高的血清胆固醇水平。PCSK与动脉粥样硬化的关系除了ApoE和LDLR基因外,还有其他与血脂调控相关的类似于PCSK9等靶点用于构建动脉粥样硬化模型,这些模型具有不同的致病机制和病理表型,助力动脉粥样硬化和高脂血症等疾病的研究及治疗药物的开发[6]。PCSK9是近年来在降脂药开发领域研究的一个新兴靶点,主要在肝脏中表达。PCSK9在LDL-C代谢中发挥重要作用。在肝细胞质膜,分泌的PCSK9的催化结构域与LDL-R结合并被内化,进入内体途径。内体的低pH值增强了PCSK9对LDL-R的亲和力,防止受体再循环到细胞表面。而PCSK9与LDL-R结合后再与LDL-C结合形成新的复合体进入溶酶体中一起被降解,最终导致LDL-R 减少,LDL-C的降解随之减少,从而使LDL-C的水平升高[7]。 PCSK9 在 LDL 受体 (LDL-R) 代谢中的作用[7]脂质代谢分析B-hPCSK9 小鼠的脂质代谢分析。分析 B-hPCSK9 小鼠和野生型 C57BL/6 小鼠(n=36,6周)TG、TC、LDL-C和 HDL-C 的血浆浓度。B-hPCSK9小鼠和野生型 C57BL/6 小鼠之间无差异。TG:甘油三酯;TC:总胆固醇;HDL-C:高密度脂蛋白胆固醇;LDL-C:低密度脂蛋白胆固醇。药效验证抗人 PCSK9 抗体在 WD 诱导 B-hPCSK9 小鼠的体内有效性。B-hPCSK9 小鼠给予Alirocumab(内部)/Evolocumab(内部)或同型对照抗体(单次给药,s.c.)(n=8,雄鼠)。在第-5、1、3、5和8天采集血液进行分析。与同型对照相比,抗人 PCSK9 抗体给药组小鼠的血清 LDL-C(A) 和 TC(B) 水平降低,表明抗人 PCSK9 抗体可有效控制雄性 B-hPCSK9 小鼠的血脂。数值表示为平均值±SEM。TC:总胆固醇;LDL-C:低密度脂蛋白胆固醇。WD:西方饮食。抗人 PCSK9 抗体上调 B-hPCSK9 小鼠的 LDLR 水平。B-hPCSK9 小鼠给予Alirocumab(内部)/Evolocumab(内部)或同型对照抗体(单次给药,s.c.)(n=6,雄鼠)。在第8天采集肝组织用于 ELISA 分析。与同种型对照相比,抗人 PCSK9 抗体给药组小鼠中 LDLR 水平上调。数值表示为平均值±SEM。LDLR:低密度脂蛋白胆固醇受体。WD:西方饮食。表1. 动脉粥样硬化相关小鼠模型参考文献[1]Besa Emini Veseli,Paola PerrottaEuropean Journal of Pharmacology,Animal models of atherosclerosis,Volume 816, 5 December 2017.[2]Wikipedia/CC BY 3.0[3]Godfrey, S, Getz, Catherine, A, & Reardon. (2012). Animal models of atherosclerosis. Arteriosclerosis Thrombosis & Vascular Biology.[4]Jean Davignon, Jeffrey S.Cohn, et al. Apolipoprotein E and atherosclerosis: insight from animal and human studies.Clinica Chimica Acta.Clinica Chimica Acta.[5]Brown, Michael S, and Joseph L. Goldstein. A receptor-mediated pathway for cholesterol homeostasis. Science232.4746(1986): 34-47.[6]Gresham, G. A. (2017). Animal models of atherosclerosis. Biomedical Reports, 6(3), 259-266.[7]Hess, C. N., Low Wang, C. C. & Hiatt, W. R. PCSK9 Inhibitors: Mechanisms of Action, Metabolic Effects, and Clinical Outcomes. Annual review of medicine 69, 133-145

随着生活水平的提高以及医疗技术的不断发展,人类预期寿命在逐步延长,这导致全球老年人口的空前增长,与此同时,与年龄相关的疾病负担也呈指数级增长[1]。有研究指出,在健康状况没有得到改善的情况下单纯延长寿命,是没有意义的[2]。基于此,世界卫生组织在2015年提出了《老龄化与健康全球战略和行动计划》,旨在实现健康老龄化的目标[3]。为了实现这一目标,需要对生物体、器官和细胞水平的衰老及疾病过程有透彻的了解,并针对衰老相关的分子途径开发预防和治疗衰老相关疾病的策略[4]。我国65岁以上人口占总人口比重及预测[5]大多数自发肿瘤、代谢性疾病以及神经退行性疾病的发病率和流行率随年龄增长而增加,其主要原因可以归咎于老年人免疫系统失调。免疫系统可以抵御病原体和杀伤癌细胞。老年人的免疫系统老化,使得免疫反应性急剧下降并且功能失调,这一现象被称为免疫衰老。研究表明,免疫治疗干预在幼小动物身上可能是有效的,但同样的治疗方法在老年动物身上却并不一定有效。考虑到大多数的癌症患者是老年人,癌症的发病率随着老龄化而增加,因此理解免疫系统失调对于免疫治疗的影响,可以更好的干预老年人的癌症治疗进程。为了最优地激发老年人的抗肿瘤免疫反应,需要识别和了解老年人免疫系统的内在缺陷,并使用自身耐受和衰老同时存在的相关模型。[6]小鼠凭借其易于饲养和处理并且寿命较短(~2.5年)的特性,成为衰老及其相关疾病研究中有吸引力的哺乳动物模式生物。[7]百奥动物提供优质老龄鼠(目前最大可提供15月龄,即60周龄的小鼠,陆续可提供最大20-22月龄小鼠),可用于衰老及其相关疾病的研究。可提供品系如下:老龄鼠百奥动物老龄鼠优势① 不同周龄小鼠现货提供;② 高标准的饲养环境;③ 非生产退役种鼠;④ 严格的遗传稳定性监测。C57BL/6J老龄鼠C57BL/6J是使用最为广泛的近交品系之一,也是第一个完成基因组测序,特征分析最全面的小鼠品系。有研究人员对C57BL/6J的生存曲线进行分析,并提供了可比较的对应的人类年龄。衰老是个连续的动态过程,大多数研究将生命阶段简单的分为成年、中年和老年。小鼠月龄和人类年龄对照关系及特点① 小鼠的3-6月龄对应成年人(20-30岁);发育完成尚未受到衰老影响。② 小鼠的10-15月龄对应中年人(38-47岁);可以检测到部分衰老生物标志物(非全部)的衰老变化。③ 小鼠的18-24月龄对应老年人(56-69岁);可以检测到几乎全部衰老标志物的衰老变化[8]。老龄鼠常见表型及病理随着小鼠生长发育,外观会逐渐改变。一些常见的衰老表型有:眼周脱毛、嘴鼻部被理毛、背侧产生脱毛区及灰毛区等。C57BL/6J老龄鼠的部分正常衰老表型[9]同时随着年龄增长,与年龄相关的各项疾病也随之而来,产生一些和疾病相关的表型。例如:自发肿瘤、眼周病变、皮炎脱毛等。非近亲繁殖小鼠长期研究(2年或更长时间)的常见疾病[10]a、Findings from multiple studies are summarized for each line or group. and original terminology.b、4-way crossc、(Af _ C57BL/6)F1d、UM HET3 from (BALB/cByJ _ C57BL/6J)F1 _ (C3H/HeJ _ DBA/2J)F1 老龄鼠应用研究① 健康生命周期的基础生物学研究② 骨质、肌肉、身体组成相关研究③ 饮食、运动、认知与行为研究④ 心血管、内分泌、代谢等疾病研究⑤ 免疫学、肿瘤学、神经退行性疾病研究⑥ 年龄相关的视力、听力障碍研究百奥动物自主研发人源化小鼠模型、疾病模型、老龄鼠等上千种小鼠模型,欢迎来电咨询~参考文献:[1] Olshansky, S.J. From Lifespan to Healthspan. Jama 320, 1323-1324 (2018).[2] Rivera-Tavarez, C.E. Can We Increase Our Health Span? Physical medicine and rehabilitation clinics of North America 28, 681-692 (2017).[3] Beard, J.R. et al. The World report on ageing and health: a policy framework for healthy ageing. Lancet (London, England) 387, 2145-2154 (2016). [4] Barzilai, N., Cuervo, A.M. & Austad, S. Aging as a Biological Target for Prevention and Therapy. Jama 320, 1321-1322 (2018).[5]http://news.youth.cn/jsxw/202103/t20210310_12758403.htm[6] J. Lustgarten.Cancer, aging and immunotherapy: lessons learned from animal models.Cancer Immunol Immunother 58:1979–1989(2009) .[7] Mitchell, S.J., Scheibye-Knudsen, M., Longo, D.L. & de Cabo, R. Animal models of aging research: implications for human aging and age-related diseases. Annual review of animal biosciences 3, 283-303 (2015). [8] Flurkey, K., M. Currer, J. & Harrison, D.E. Chapter 20 - Mouse Models in Aging Research. In: Fox, J.G. et al. (eds). The Mouse in Biomedical Research (Second Edition). Academic Press: Burlington pp 637-672(2007).[9] https:// e. dxy. cn/ broadcast/ live/ id/ 22788/ ? source= wechat& sim= 1& sim_ token= jo3ALEk4m RZ9tfoghOHA rih1tV 6kp0IQvMAdhr7vws7ey9YqHi8uJxdmTk9vElaf[10] C. F. Brayton, P. M. Treuting, and J. M. Ward.Pathobiology of Aging Mice and GEM: Background Strains and Experimental Design.Veterinary Pathology 49(1) 85-105(2012).

1973年,Kobylka和Carraway在乳腺上皮细胞中发现了一种在乳脂球中无法水解的膜蛋白,1978年,该分子被鉴定为血小板GPIV,一种介导血小板中血小板反应蛋白-1(TSP-1)结合的蛋白。随后,Tandon发现GPIV与白细胞分化抗原CD36具有重叠结构。随后,其他研究人员确定了CD36在血小板活化和细胞粘附中的作用。1993年,Abumrad及其同事证明了CD36的脂肪活性,之后将该蛋白在小鼠体内克隆并鉴定为人CD36的小鼠直系同源物,表明其在脂肪酸摄取中的作用。CD36现在被广泛认为是一种清道夫受体,可以吸收长链脂肪酸(LCFAs)和氧化低密度脂蛋白(ox-LDL)。CD36是一种清道夫受体,在多种细胞类型中表达。在脂质代谢中发挥重要作用,介导脂质摄取、免疫识别、炎症、分子粘附和凋亡。与血管生成、炎症反应、动脉粥样硬化性血栓性疾病以及代谢紊乱性糖尿病和肥胖症等有关。在免疫系统中,CD36介导树突状细胞抗原的获取和呈递,并支持调节性T细胞功能。研究表明靶向CD36可能是提高CD8+T细胞免疫疗法的抗肿瘤药效的有效策略,为临床治疗提供了新的思路和可能。[1]CD36与PPAR-β激活[2]CD36靶向作用破坏肿瘤内Treg细胞,并启动PD-1阻断肿瘤。具有抗CD36抗体的MOA: ①耗尽CD36+ Treg细胞。②阻断PPAR-和PPAR-依赖的代谢途径,降低线粒体活性和CD36表达水平,促进Treg细胞凋亡。③阻断TSP-1/CD36通路,促进TME血管生成。CD36调节肿瘤免疫[3]CD36在肿瘤微环境中发挥作用。CD36调节下游Src家族激酶促进抗血管生成、FAO以及化疗耐药和放疗耐药,从而导致肿瘤转移。CD36还通过EMT激活Wnt/TGF-β信号传导促进肿瘤转移。CD36摄取多种脂质,如ox-LDL、LCFAs和胆固醇。免疫细胞中的脂质沉积导致树突状细胞的无菌性炎症和抗原呈递功能障碍,从而诱导肿瘤免疫抑制。此外,CD36可以与凋亡细胞结合并激活交叉引物,这可能导致免疫抑制和肿瘤发展。BioMice百奥动物自主研发了人源化小鼠B-hCD36 mice和人源化细胞系B-hCD36 MC38助力科研。B-hCD36 miceB-hCD36小鼠血常规检测全血细胞计数(CBC)。采集雌性C57BL/6和B-hCD36小鼠(n=8,6-8周龄)的血液并进行全血细胞计数分析。B-hCD36小鼠的测量结果与C57BL/6小鼠相似,表明人源化不会改变血细胞组成和形态。数值表示为平均值±SEM。B-hCD36小鼠血生化检测B-hCD36小鼠的血生化试验。收集C57BL/6和B-hCD36小鼠(n=8,6-8周龄)的血清,并分析其指标水平。B-hCD36小鼠的测量结果与C57BL/6小鼠相似,表明人源化不会改变肝脏等相关组织的健康。数值表示为平均值±SEM。使用抗CD36抗体测试B-hCD36模型的抗体结合试验通过流式细胞术对纯合B-hCD36小鼠进行种属特异性CD36表达分析。收集野生型小鼠 (+/+)和纯合B-hCD36小鼠(H/H)的腹腔渗出巨噬细胞,并通过抗CD36抗体的流式细胞术进行分析。人CD36仅在纯合B-hCD36小鼠中可检测到,但在野生型小鼠中未检测到。CD36-1G04-hIgG1-LALA抗体在小鼠和人中交叉识别。B-hCD36 MC38蛋白表达分析通过流式细胞术对B-hCD36 MC38细胞中的CD36表达进行分析。用种属特异性抗 CD36抗体对B-hCD36 MC38培养物的单细胞混悬液进行染色。在B-hCD36 MC38细胞表面检测到人CD36,但在野生型MC38细胞表面未检测到。使用B-hCD36 MC38细胞的10-G12克隆进行体内实验。肿瘤生长曲线和体重变化B-hCD36 MC38细胞的皮下同种移植肿瘤生长。将B-hCD36 MC38细胞(5x105)和野生型MC38细胞(5x105)皮下植入B-hCD36小鼠(雌性,7周龄,n=5)。每周测量两次肿瘤体积和体重。(A)平均肿瘤体积±SEM。(B)体重(平均值±SEM)。体积以mm3表示,使用公式:V=0.5X长径X短径2。如图A所示,B-hCD36 MC38细胞能够在体内建立肿瘤,可用于有效性研究。肿瘤细胞蛋白表达分析将B-hCD36 MC38细胞皮下移植到B-hCD36小鼠(n=5)中,接种后35天,收获肿瘤细胞并通过流式细胞术评估人CD36表达。如图所示,人CD36在肿瘤细胞表面高度表达。因此,B-hCD36 MC38细胞可用于新型CD36疗法的体内药效研究。更多验证数据信息,正在研究进行中,敬请关注。想要获取更多百奥动物自主研发的人源化小鼠信息,请访问百奥动物官网或扫描下方二维码。参考文献1.CD36-mediated ferroptosis dampens intratumoral CD8+T cell effector function and impairs their antitumor ability2.Wang, H., Franco, F., Tsui, Y. et al. CD36-mediated metabolic adaptation supports regulatory T cell survival and function in tumors. Nat Immunol 21, 298–308 (2020). doi:10.1038/s41590-019-0589-53.Wang, Jingchun; Li, Yongsheng (2019). CD36 tango in cancer: signaling pathways and functions. Theranostics, 9(17), 4893–4908. doi:10.7150/thno.36037
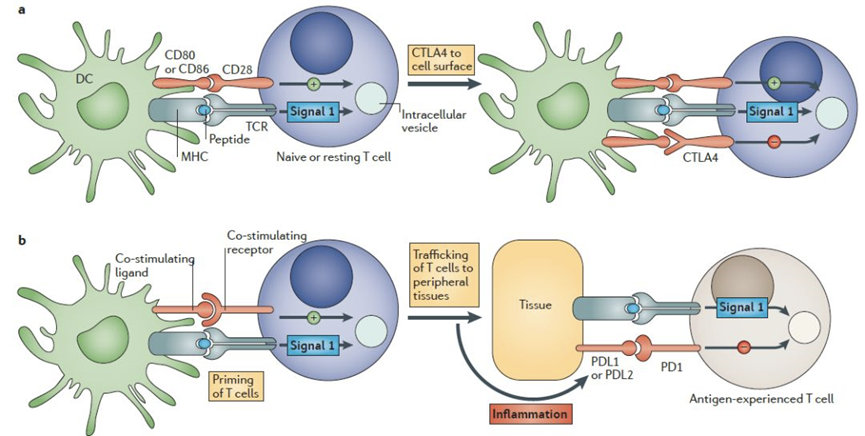
2006年3月,TeGenero公司的CD28超级激动剂单抗TGN1412治疗风湿性关节炎/白血病的1期临床试验中,6名志愿者遭受细胞因子释放综合征并伴有多器官功能衰竭,全身极度肿胀成为“大象人”,万幸的是最终脱离生命危险,该事件直接导致TeGenero破产。16年前的黑天鹅事件也给CD28药物开发蒙上了一层阴影。后续TGN1412并未被就此放弃,莫斯科抗体公司TheraMab顺利接盘并更名为TAB08,有报道称1期临床NCT03006029、NCT01990157不良事件保持在一个可接受的水平(短暂的发热与IL-6有关),然而试验如今已经因“行政原因”终止,这款CD28单抗最终也没能迎来涅槃重生[1]。如此看来,再次沉沦的CD28的未来当真就是砂砾宿命了吗?2017年,Science上发表了一篇题为Rescue of exhausted CD8 Tcells by PD-1 targeted therapies is CD28-dependent的文章,证明了CD28/B7共刺激途径对有效的PD-1疗法至关重要。至此,CD28再次回到研究者的视线中。2019年11月,赛诺菲在Nature Cancer发表了一款CD28/CD3/CD38三抗的研究进展。该三抗在CD3分子亲和力的选择上,采用了中等亲和力的抗体(KD~20nM)平衡了杀伤有效性以及细胞因子大量释放的安全性问题[2]。2020年1月,再生元在Science Translational Medicine发表论文,指出CD28双抗可以增强CD3的抗肿瘤疗效。在动物实验中,共刺激型CD28双抗明显增强了CD3双抗的疗效,且没有细胞因子风暴的风险。2020年6月24日,再生元再次在ScienceTranslational Medicine杂志上发表论文,证实了肿瘤特异性抗原(TSA) x CD28双特异性抗体可以与更广泛的抗PD-1抗体协同作用,增强癌症治疗效果,诱导长效抗肿瘤免疫力,且不会诱发细胞因子风暴,具有很好的耐受性。今年2月,赛诺菲的研发团队在Nature发表了一篇HER2×CD3×CD28三抗的研究进展。研究表明,在原代人CD3+T细胞重组的免疫缺陷NSG小鼠体内,该三抗使CD8 T细胞中颗粒酶的表达增加了6.8倍。相关肿瘤药效实验结果表明,剂量低至10μg/kg时,HER2×CD3×CD28三抗依然能够诱导肿瘤消退。近期,惠和生物靶向CD3×CD28×CD19三特异性抗体CC312的IND申请获得美国FDA默示许可,进入临床研发阶段,CC312是国内首个,也是全球第三个基于CD28共刺激信号的三特异性抗体。随着赛诺菲、再生元等牵头的研究推进,CD28被强势拉回公众视野,更多的潜力有待开发!CD28相关信号通路CD28是T细胞激活最重要的参与者之一,是增强MHC-TCR的主激活信号的主要辅助信号。CD28和CTLA-4具有高度的同源性,具有相同的配体CD80和CD86(B7-1和B7-2),CD28负责传递激活信号,活化T细胞,而CTLA-4负责传递抑制信号给T细胞,让T细胞不会杀伤其它细胞,包括肿瘤细胞。CTLA-4与配体的亲和力要高于CD28,因此会竞争阻断CD28的T细胞激活作用。在T细胞辅助激活的过程中,CD86优先表达,在与CD28结合上,CD86表现出优于CD80的结合能力,有利于T细胞激活。在T细胞激活的过程中,CD28扮演着“加油”角色,CTLA-4扮演着“前刹车”角色,PD-1则扮演着“后刹车”角色[3]。CD28、CTLA-4、PD-1/PD-L1信号通路[4]CD28靶点部分开发进展CD28作为一个老靶点,不仅机制清晰,而且针对不同临床适应症,既可开发拮抗剂也可开发激动剂,理应前景无限,但由于其研发历程的跌宕起伏,目前只有少量抗体药物开发管线。其临床药物开发上主要集中在3个方面:CD28关联靶点融合蛋白药物;单抗药物;多特异性抗体药物。(数据来源科睿唯安及公开信息整理)CD28系列人源化动物模型对于CD28相关药物的研究,再生元和赛诺菲已经做出了很好的表率,进一步的数据也许会让CD28再次火热起来。相关动物模型对于CD28靶向调节剂开发可谓至关重要,BioMice 百奥动物自主研发的CD28系列人源化鼠是评估CD28相关抗体药物的优质临床前实验动物模型。B-hCD28 小鼠验证数据蛋白表达分析流式细胞术分析B-hCD28纯合小鼠CD28的表达取抗CD3ε抗体刺激野生型小鼠和纯合B-hCD28小鼠的脾细胞,用种属特异性抗CD28抗体进行流式分析。小鼠CD28在野生型小鼠中检测到。人CD28仅在纯合B-hCD28小鼠中检测到,而在野生型小鼠中检测不到。抗人CD28抗体的体内药效抗人CD28抗体在B-hCD28小鼠体内的抗肿瘤活性实验结果表明,两种抗人CD28抗体在B-hCD28小鼠体内均能有效控制肿瘤生长,表明B-hCD28小鼠为抗人CD28抗体的体内评价提供了一个优质的临床前模型。抗人PD-L1xCD28双特异性抗体的体内药效抗PD-L1和抗CD28 (PD-L1xCD28)双特异性抗体(BsAb)在B-hCD28小鼠中的抗肿瘤活性结果表明,抗人PD-L1xCD28 BsAbs能够有效控制B-hCD28小鼠的肿瘤生长,表明B-hCD28小鼠为抗人PD-L1xCD28 BsAbs的体内评价提供了一个优质的临床前模型。B-hCD3E/hCD28小鼠验证数据蛋白表达分析流式细胞术分析B-hCD3E/hCD28纯合小鼠CD3E的表达取野生型小鼠和纯合B-hCD3E/hCD28小鼠脾细胞,用种属特异性抗CD3E抗体进行流式细胞术分析。小鼠CD3E在野生型小鼠中检测到。人CD3E仅在纯合B-hCD3E/hCD28小鼠中检测到,而在野生型小鼠中检测不到。流式细胞术分析B-hCD3E/hCD28纯合小鼠CD28的表达取野生型小鼠和纯合B-hCD3E/hCD28小鼠脾细胞,用种属特异性抗CD28抗体进行流式细胞术分析。小鼠CD28在野生型小鼠中检测到。人CD28仅在纯合B-hCD3E/hCD28小鼠中检测到,而在野生型小鼠中检测不到。B-hCD3EDG/hCD28 小鼠验证数据蛋白表达分析流式细胞术分析B-hCD3EDG/hCD28纯合小鼠CD3E的表达取野生型小鼠和纯合B-hCD3EDG/hCD28小鼠脾细胞,用种属特异性抗CD3E抗体进行流式分析。小鼠CD3E在野生型小鼠中检测到。人CD3E仅在纯合B-hCD3EDG/hCD28小鼠中检测到,而在野生型小鼠中检测不到。流式细胞术分析B-hCD3EDG/hCD28纯合小鼠CD28的表达取野生型小鼠和纯合B-hCD3EDG/hCD28小鼠脾细胞,用种属特异性抗CD28抗体进行流式细胞术分析。小鼠CD28在野生型小鼠中检测到。人CD28仅在纯合B-hCD3EDG/hCD28小鼠中检测到,而在野生型小鼠中检测不到。B-hCD28/hTROP2小鼠验证数据蛋白表达分析流式细胞术分析B-hCD28/hTROP2纯合小鼠CD28的表达取野生型小鼠和纯合B-hCD28/hTROP2小鼠脾细胞,用种属特异性抗CD28抗体进行流式细胞术分析。小鼠CD28在野生型小鼠中检测到。人CD28仅在纯合B-hCD28/hTROP2小鼠中检测到,而在野生型小鼠中检测不到。Western blot分析TROP2在B-hCD28/hTROP2纯合小鼠中的表达取野生型小鼠和纯合B-hCD28/hTROP2小鼠的皮肤组织,用抗TROP2抗体进行western blot分析。小鼠TROP2在野生型小鼠中检测到。人TROP2仅在纯合B-hCD28/hTROP2小鼠中检测到,而在野生型小鼠中检测不到。B-hCD28/hB7-H3小鼠验证数据蛋白表达分析流式细胞术分析B-hCD28/hB7-H3纯合小鼠CD28的表达取野生型小鼠和纯合B-hCD28/hB7-H3小鼠脾细胞,用种属特异性抗CD28抗体进行流式细胞术分析。小鼠CD28在野生型小鼠中检测到。人CD28仅在纯合B-hCD28/hB7-H3小鼠中检测到,而在野生型小鼠中检测不到。Western blot分析B-hCD28/hB7-H3纯合小鼠B7-H3的表达采集野生型小鼠和纯合B-hCD28/hB7-H3小鼠附睾,用抗B7-H3抗体进行western blot分析。由于抗体的交叉反应,野生型小鼠和纯合B-hCD28/hB7-H3小鼠均可检测到B7-H3。B-hCD28/hCD20小鼠验证数据蛋白表达分析流式细胞术分析B-hCD28/hCD20纯合小鼠CD28的表达取野生型小鼠和纯合B-hCD28/hCD20小鼠脾细胞,用种属特异性抗CD28抗体进行流式细胞术分析。小鼠CD28在野生型小鼠中检测到。人CD28仅在纯合B-hCD28/hCD20小鼠中检测到,而在野生型小鼠中检测不到。流式细胞术检测B-hCD28/hCD20纯合小鼠CD20的表达取野生型小鼠和纯合B-hCD28/hCD20 小鼠脾细胞,用种属特异性抗CD20抗体进行流式细胞术分析。小鼠CD20在野生型小鼠中检测到。人CD20仅在纯合B-hCD28/hCD20小鼠中检测到,而在野生型小鼠中检测不到。B-hSIRPA/hCD47/hCD3E/hCD28小鼠验证数据蛋白表达分析流式细胞术分析B-hSIRPA/hCD47/hCD3E/hCD28纯合小鼠CD3E、CD28、SIRPA、CD47的表达取野生型小鼠和纯合B-hSIRPA/hCD47/hCD3E/hCD28小鼠脾细胞,分别用种特异性抗CD3E、CD28、SIRPA、CD47抗体进行流式细胞术分析。由于抗体的交叉反应,野生型小鼠和B-hSIRPA/hCD47/hCD3E/hCD28小鼠均可检测到小鼠SIRPA,人SIRPA仅在纯合B-hSIRPA/hCD47/hCD3E/hCD28小鼠中检测到,而在野生型小鼠中未检测到;小鼠CD3E、CD28、CD47在野生型小鼠中检测到,人CD3E、CD28、CD47仅在纯合B-hSIRPA/hCD47/hCD3E/hCD28小鼠中检测到,而在野生型小鼠中检测不到。CD28靶点相关模型列表 想要了解更多人源化小鼠,欢迎扫描下方二维码查看或者来电咨询。 参考资料:[1] https://mp.weixin.qq.com/s/6tuPYtTIr_gNQrb0N7o7_w[2] Sanofi R&D Investor Event[3] Dimitris Skokos et al. A class of costimulatoryCD28-bispecific antibodies that enhance the antitumor activity ofCD3-bispecific antibodies. Sci. Transl. Med. 2020[4]https://oncologypro.esmo.org/education-library/esmo-handbooks/immuno-oncology/immune-synapse
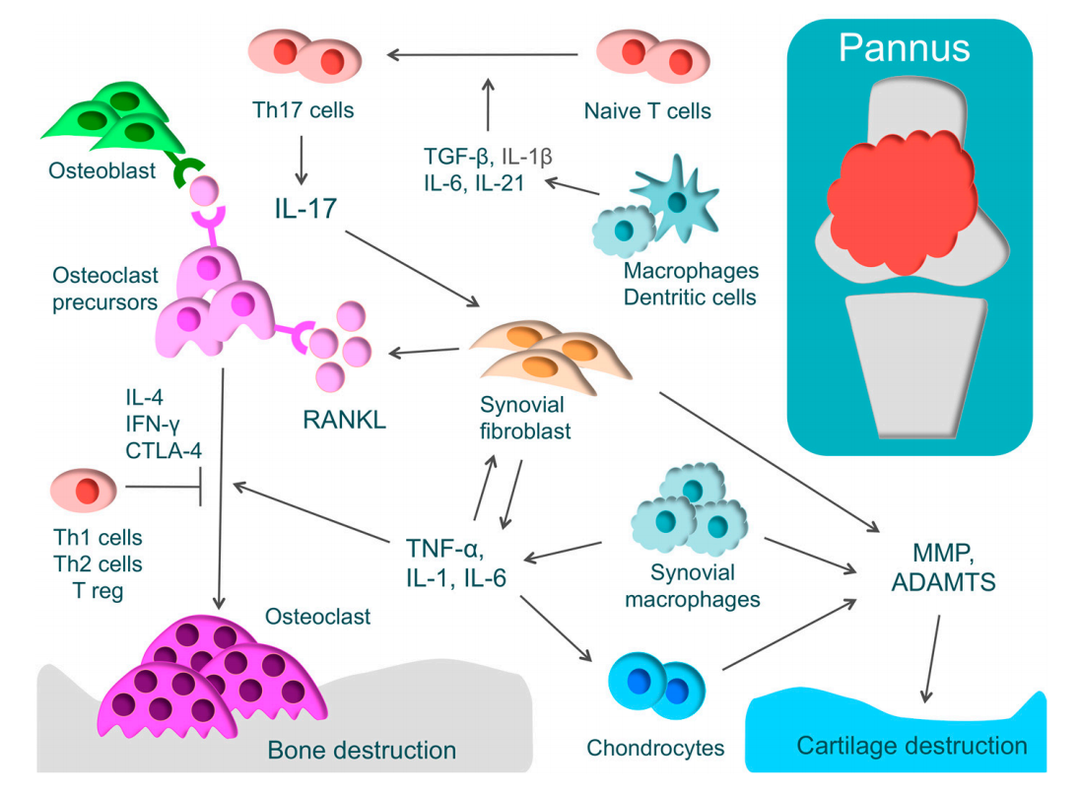
不死的癌症类风湿性关节炎(Rheumatoid Arthritis,RA)是人类最常见的自身免疫性疾病之一,其临床症状主要表现有全身关节肿胀、疼痛和运动失调等,病情极其严重的甚至可能会导致残疾。而病理主要表现出增生性滑膜炎,软骨损伤以及骨结构的破坏。为了更好的研究人类的类风湿性关节炎,研究者在小鼠上开发出多种能模拟人类类风湿性关节炎的动物模型,而这之中胶原诱导型关节炎,能最大程度反映出人类类风湿性关节炎的临床和病理特征。Rheumatoid Arthritis百奥动物在不同品系的小鼠(C57BL/6、DBA)上都建立了稳定的胶原诱导型关节炎疾病模型(Collagen-Induced Arthritis,CIA),可用于相关药物的药效评价。类风湿性关节炎 (RA) 的病理及软骨和骨破坏的机制RA 的特征是增生性滑膜(血管翳)和 T 细胞的过度免疫反应。血管翳包括 T 细胞、滑膜成纤维细胞和巨噬细胞,可产生炎性细胞因子,如肿瘤坏死因子 (TNF)-α、白细胞介素-1(IL-1)、IL-6和IL-17。相关数据展示01RA/CIA 模型介绍 临床评分利用胶原(CII)在C57BL/6小鼠中诱导关节炎模型。(A)小鼠体重变化;(B)临床评分;(C)小鼠发病率。结果显示:在造模组(G2)中,小鼠的临床评分显著增加,提示关节炎模型建立成功。 病理分析C57BL/6小鼠关节炎模型建立后的病理分析。(A)病理评分;(B)病理切片H&E染色。造模组动物全部或部分关节可见皮下混合炎细胞浸润(a),关节周围狭窄(b),踝关节和/或趾骨关节软骨和骨组织破坏(c)等关节炎病变,进一步提示关节炎模型建立成功。02C57BL/6小鼠关节炎模型用于评价甲氨蝶呤(MTX)药效 临床评分甲氨蝶呤在小鼠关节炎模型中的药效作用。(A)小鼠体重变化;(B)临床评分;(C)小鼠发病率。结果显示:在造模组(G2)中,小鼠的临床评分显著增加,表明CIA模型建立成功,同时相比于未造模组(G1),体重波动更加明显。在给于甲氨喋呤(MTX)后,治疗组(G3)的平均临床评分显著低于造模组,证明小鼠的病情得到了有效控制。另外,在发病率方面,治疗组的发病率最大值为40%,也明显低于造模组近80%的发病率,提示小分子药甲氨喋呤对该疾病具有治疗作用。 病理分析甲氨喋呤在小鼠关节炎模型中药效作用的病理分析。(A)病理评分;(B)病理切片H&E染色。病理结果显示:未造模组(G1)动物镜下未见明显异常改变,踝关节软骨表面光滑,关节腔明显(a)。造模组(G2)动物踝关节周围组织,可见皮下混合炎细胞浸润,关节滑膜炎和/或血管翳形成(c),关节软骨破坏,关节腔消失,部分骨组织融合(b)。与G2造模组相比,治疗组虽然有部分踝关节周围皮下组织水肿和炎细胞浸润(d),但其病理评分均值明显低于G2组,甲氨喋呤对动物关节炎病变具有治疗作用。03B-hIL6/hIL6R小鼠关节炎模型用于评价抗人IL6抗体sirukumab药效 临床评分抗人IL-6抗体sirukumab(内部合成)在小鼠关节炎模型中药效作用。(A)小鼠体重变化;(B)临床评分。结果显示:在模型成功建立后,治疗组(G3,G4)小鼠用sirukumab抗体进行治疗,显示出治疗作用,且表现出剂量依赖性。 病理分析抗人IL-6抗体sirukumab(内部合成)在小鼠关节炎模型中药效作用的病理分析。(A)病理切片H&E染色;(B)病理评分。结果显示:未造模组(G1)动物未见明显异常改变,踝关节软骨表面光滑,关节腔明显。造模组(G2)动物踝关节骨组织损伤(d),关节腔或关节周围腔隙消失(e),并出现血管翳(a)。与造模组相比,抗体药治疗低剂量组(G3)有部分炎细胞浸润(b)和血管翳(a)生成,并表现出滑膜增生(c)。但在抗体药治疗高剂量组(G4),仅有部分血管翳,关节炎病变基本消失,关节腔明显,提示sirukumab对小鼠关节炎具有治疗作用。04服务项目更多药效相关服务,CAIA引起的关节炎模型,骨关节炎模型,骨质疏松模型,正在研究进行中,感兴趣的欢迎联系我们获取资料。想要获取更多百奥动物自主研发的自身免疫性疾病模型信息,请访问百奥动物官网。参考文献:1. Tateiwa, D., Yoshikawa, H. & Kaito, T. Cartilage and Bone Destruction in Arthritis: Pathogenesis and Treatment Strategy: A Literature Review. Cells 8, doi:10.3390/cells8080818 (2019).2. McInnes IB, et al. Cytokines in the pathogenesis of rheumatoid arthritis. Nat Rev Immunol 7, 429-42 (2007).
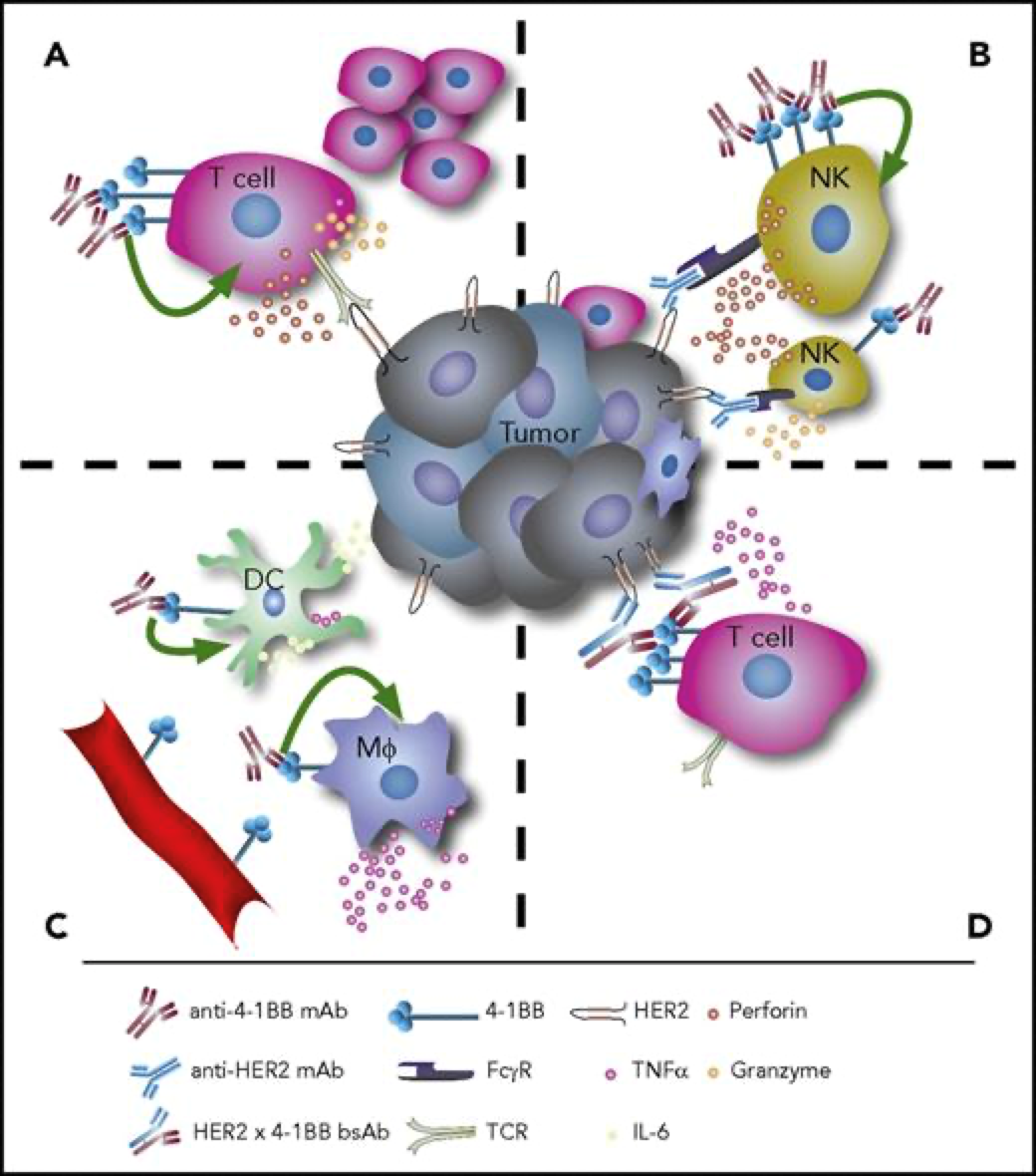
4-1BB靶点介绍4-1BB,即CD137,TNFRSF9(TNF receptor superfamily member 9),它于1989年首次被发现,属于TNF受体超家族。它在抗原启动的T细胞上表达,在静止的T细胞上不表达,除此以外,4-1BB还在树突状细胞(DC)、自然杀伤细胞(NKs)、活化的CD4+和CD8+T淋巴细胞、嗜酸性粒细胞、自然杀伤T细胞(NKT)和肥大细胞中表达。当4-1BB与其配体4-1BBL结合后,可以产生共刺激信号诱导CD4+和CD8+T细胞的活性,促进T细胞增殖,刺激巨噬细胞产生如如IL-6、TNF-α等多种炎症细胞因子。从而激活机体免疫系统,达到抑制肿瘤发生发展、促进肿瘤细胞清除的功效。有研究报道,当在进行4-1BB抗体药物研发时,还要需要考虑抗体结合位点在人和小鼠之间存在的差异性。其结合作用仅限于富含半胱氨酸结构域(CRD),小鼠4-1BBL主要与CRDII结合,人4-1BBL主要与CRDIII结合。小鼠4-1BB/4-1BBL是二聚体,人4-1BB/4-1BBL为三聚体,且人4-1BB不与小鼠的4-1BBL相互作用。[1]图1. 抗4-1BB疗法的免疫调节作用[2]4-1BB药物开发的现状4-1BB作为癌症免疫疗法中十分有潜力的靶点,针对其进行的药物研发非常火热,全球目前已有至少40款4-1BB激动型抗体进入临床研究阶段。4-1BB单抗药物由 BMS研发的Urelumab(全人源IgG4 mAb)是一个4-1BB强激动性的单抗,在I/II期试验中表现出良好的抗肿瘤活性,但在后续治疗过程中发生了两起由肝毒性引起的致命性不良事件。由Pfizer研发的Utomilumab(人源化IgG2 mAb)是一个4-1BB弱激动性的单抗,虽然有较好的肝脏安全性表现,但作为单药治疗抗肿瘤活性十分有限。因此,此前未能很好的在激动作用和肝毒性之间达到良好的平衡,导致4-1BB单抗产品临床应用空间有限;目前新一代单抗设计通过对Fc功能改造试图保留激动性的同时克服安全性问题,Alligator、Agenus以及国内的礼进生物和天演药业的4-1BB单抗产品表现出良好的初步临床数据。4-1BB重点单抗产品临床进度数据来源:Cortellis4-1BB双抗药物平衡抗体激动作用强弱和其肝毒性,是4-1BB单抗药物开发的难点,因此为了克服疗效和毒性的问题,研究者们开始探索新的开发策略。其中以4-1BB为靶点的双抗、三抗以及四抗成为研发的一个热点领域,国内外多家药企均有布局。4-1BB双特异性抗体通过对肿瘤细胞表面靶点和4-1BB亲和力不同精准定位肿瘤微环境,减少肝毒性等副作用。因此双抗疗效相比单抗可以实现更精准的靶向,从而使得临床前的抗肿瘤活性和安全性都得到了提升。如PD-1/PD-L1就是4-1BB双抗常用的另一候选靶点。去年,BioNTech与Genmab联合研发的PD-L1×4-BB双抗药物GEN1046,是一种全长IgG1亚型的PD-L1×4-1BB双抗,其以临床Ⅰ/Ⅱa期结果发表在著名学术期刊《CANCER DISCOVERY》上的文章中,所使用的PD-L1/4-1BB双靶点人源化鼠由百奥动物提供。文中研究结果表明:GEN1046对于CPI耐药的实体瘤治疗效果良好,安全性高,为免疫联合治疗提供了新的方案。4-1BB重点双抗产品临床进度数据来源:Cortellis通过众多临床试验及以往多年研究积累,4-1BB 靶点被证实是肿瘤免疫治疗十分有潜力的靶点,目前如何最大化激动作用并最小化毒性是未来4-1BB激动剂研发亟需解决的问题。百奥动物自主研发的4-1BB系列人源化小鼠(详情见下表),是靶向4-1BB抗体开发相关药物进行药效评价的优质模型。B-h4-1BB/h4-1BBL mice高剂量Urelumab对B-h4-1BB小鼠和B-h4-1BB/h4-1BBL小鼠均产生肝毒性用PBS或Urelumab(内部合成)对纯合子B-h4-1BB小鼠和B-h4-1BB/h4-1BBL小鼠进行治疗 (n=5,雌性,7周龄)。第21天采集血清,检测谷丙转氨酶(ALT)、谷草转氨酶(AST)。与PBS对照相比,使用20mg /kg Urelumab治疗B-h4-1BB小鼠的ALT显著升高,而同样的治疗对B-h4-1BB/h4-1BBL小鼠没有影响。与PBS对照相比,B-h4-1BB小鼠AST无明显升高,但20ug/mL urelumab治疗可使B-h4-1BB/h4-1BBL小鼠AST明显升高。在1mg/kg剂量组,B-h4-1BB小鼠和B-h4-1BB/h4-1BBL小鼠的ALT和AST均无明显变化。表明高剂量urelumab对B-h4-1BB和B-h4-1BB/h4-1BBL小鼠有肝毒性作用。数值以平均值±SEM表示。高剂量Urelumab 治疗可导致B-h4-1BB小鼠和B-h4-1BB/h4-1BBL小鼠肝脏淋巴细胞浸润增加肝毒性的病理分析。在B-h4-1BB小鼠(G1-G3)中,当urelumab剂量为1mg/kg时,肝脏未见明显异常变化(G2)。当剂量为20 mg/kg (G3)时,观察到血管周围细胞浸润或肝脏慢性炎症,并伴有轻度病变。在B-h4-1BB/h4-1BBL小鼠(G4-G6)中,1 mg/kg组(G5) 3/5的小鼠出现病理变化(轻度2/5,轻度1/5),而在20 mg/kg剂量(G6)时,所有实验动物肝脏均出现中度变化。总体而言,20 mg/kg组(G6)肝脏病变程度和发生率显著高于1 mg/kg组(G5)。以上结果提示,高剂量(20mg /kg)的urelumab比低剂量(1mg /kg)的urelumab更容易发生血管周围细胞浸润或肝脏慢性炎症。B-h4-1BB/h4-1BBL小鼠对urelumab毒性作用比B-h4-1BB小鼠更敏感。因此B-h4-1BB/h4-1BBL小鼠是较好的临床前毒性评价模型。B-h4-1BB/h4-1BBL药效验证抗人4-1BB抗体在B-h4-1BB/h4-1BBL小鼠中的抗肿瘤药效。(A) 纯合子B-h4-1BB/h4-1BBL小鼠 (雌性,6-8周龄,n=5) 皮下接种小鼠结肠癌MC38细胞系。结果显示抗人4-1BB抗体能够显著抑制MC38肿瘤细胞的生长。(B)治疗期间体重变化。如图A所示,抗人4-1BB抗体在B-h4-1BB/h4-1BBL小鼠中有效地控制肿瘤生长,这表明B-h4-1BB/h4-1BBL小鼠可为体内评价抗人4-1BB抗体药效提供有力的临床前研究模型。值表示为平均值±SEM。参考文献[1].https://mp.weixin.qq.com/s/kvwfybpx63XQn26xdijIZwhttps://mp.weixin.qq.com/s/999cX0RaXHLyXxZ8MhIaBw[2]. Chester C, Sanmamed MF, Wang J, Melero I. Immunotherapy targeting 4-1BB:mechanistic rationale, clinical results, and future strategies. Blood. 2018 Jan4;131(1):49-57. doi: 10.1182/blood-2017-06-741041. Epub 2017 Nov 8. PMID:29118009.
近年来,免疫治疗在抗肿瘤治疗中的作用备受关注,基于免疫治疗的新药开发和标志物探索,成为了当前肿瘤研究的热点,这也对临床前研究动物模型的建立提出了更高的要求。即可以模拟人肿瘤特征又同时存在“人源化”免疫系统的免疫重建小鼠模型,成为了免疫肿瘤研发中的优质模型。百奥动物B-NDG小鼠缺乏成熟的T、B、NK细胞,是目前国际公认的免疫缺陷程度高、适合人源细胞或组织移植的工具小鼠。将人的免疫细胞、造血干细胞移植到B-NDG及B-NDG衍生小鼠中构建的免疫系统重建小鼠,能够更好的模拟人的免疫系统,进行免疫学研究和免疫药物评价。但在使用重度免疫缺陷小鼠进行PBMC和CD34+ HSC免疫重建时会面临一些常见问题,如PBMC重建后的T细胞会对小鼠自身细胞进行攻击导致严重的xeno-GvHD反应;CD34+ HSC重建后的NK细胞和髓系细胞重建比例不足。我们以B-NDG小鼠为基础分别开发了可延缓xeno-GvHD反应和可促进髓系细胞发育的二代系列小鼠,以满足特定细胞功能研究和相应靶点药物评价的需求。减轻GvHD反应,延长实验窗口期B-NDG MHC I/II DKO mice plus 不表达MHC I/II 类分子B-NDG小鼠、B-NDG B2m KO plus小鼠和B-NDG MHC I/II DKO plus小鼠进行人PBMC重建后GvHD严重程度的比较 第0天将3个健康供体的人PBMCs (5E6)静脉注射B-NDG小鼠、B-NDG B2m KO plus小鼠和B-NDG MHC I/II DKO plus小鼠(雌性,5周龄,n=5)中。A.用Kaplan Meier生存曲线分析小鼠存活率。B.体重变化。GvHD临床体征每周评分两次。结果表明:B-NDG MHC I/II DKO plus小鼠可显著延缓GvHD发生,并降低GvHD严重程度。因此B-NDG MHC I/II DKO plus小鼠更适合将人PBMC移植到免疫缺陷小鼠模型。B-NDG B2m KO mice plus B-NDG小鼠、B-NDG B2m KO plus进行人PBMC重建后GvHD严重程度的比较将3个健康供体的2E6人PBMC静脉注射B-NDG B2m KO plus小鼠(雌性,11周龄,n=5)和B-NDG小鼠(雌性,10周龄,n=6)中。GvHD临床体征每周评分两次。结果表明:在人PBMC诱导的GvHD模型中,B-NDG B2m KO plus小鼠比B-NDG小鼠生存期明显延长,B-NDG B2m KO plus小鼠可延缓GvHD的发病并减轻其严重程度。促进髓系细胞重建B-NDG MGMT3 mice 人源化IL3, CSF2, CSF1, THPO 基因未经辐照的B-NDG MGMT3小鼠进行人CD34+HSC重建将人CD34+HSC造血干细胞(3E4)静脉注射B-NDG小鼠和B-NDG MGMT3小鼠(出生后24-72h,n=15)。B-NDG小鼠给予1.0 gy辐照,B-NDG MGMT3小鼠不辐照。流式细胞分析两种小鼠进行人CD34+HSC免疫重建后的外周血淋巴细胞。结果表明:未经辐照的B-NDG MGMT3小鼠的CD45+细胞比例从移植后12周开始达到25%,并持续上升,明显高于B-NDG小鼠。B-NDG MGMT3小鼠单核细胞、MDSCs、DCs和Treg的比例高于B-NDG小鼠。(注:A面板第18周数据因流式细胞检测问题无意义。)B-NDG hCSF1/hTHPO mice 未经辐照的B-NDG hCSF1/hTHPO小鼠进行人CD34+HSC重建将人CD34+HSC细胞(0.15 M)静脉注射到纯合B-NDG hCSF1/hTHPO小鼠(雌性,6周龄,n=15)。流式细胞术分析人CD34+HSC免疫重建后小鼠外周血淋巴细胞。结果显示:未经辐照的B-NDG hCSF1/hTHPO小鼠成功重建了T、包括B、NK、髓系细胞、单核细胞和中性粒细胞在内的人多系细胞。免疫缺陷动物产品列表
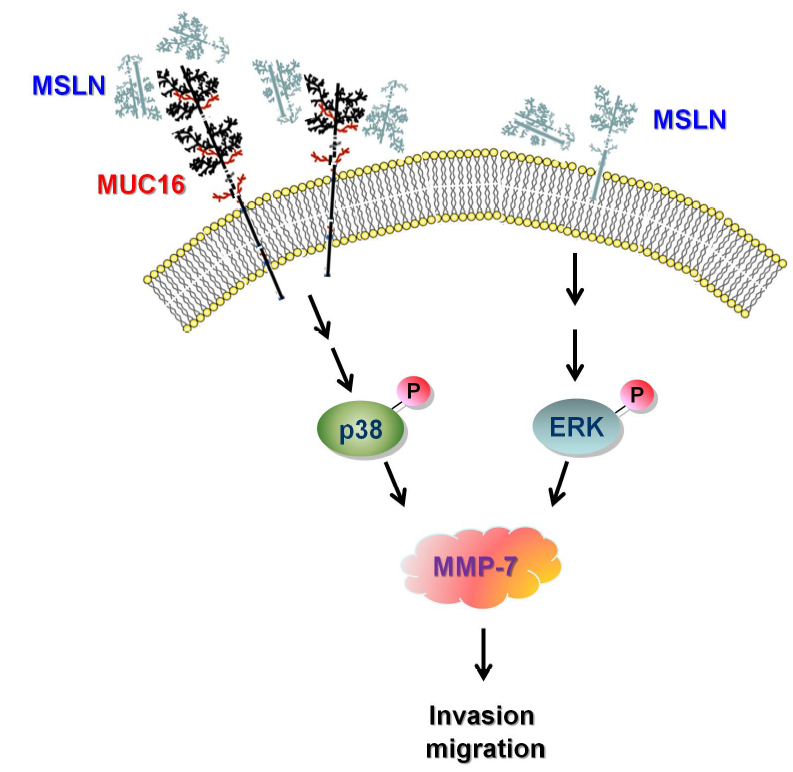
MSLN基因编码一种前体蛋白,经蛋白水解处理生成两种蛋白产物,主要位于正常间皮细胞表面,两种蛋白产物为巨核细胞增强因子(megakaryocyte potentiating,MPF)和间皮素(Mesothelin,MSLN)。巨核细胞增强因子作为细胞因子可刺激骨髓巨核细胞集落形成。间皮素在正常组织中,仅表达在间皮细胞,但在间皮瘤(mesothelioma)、卵巢癌(ovarian cancer)、胰腺癌(pancreatic cancer)、胆管癌(cholangiocarcinoma)等肿瘤中均有高表达。因此间皮素是很有前途的肿瘤特异性治疗候选药物靶点。图1.间皮素的结构和功能[1]MSLN通过ERK和PI3K/Akt通路促进肿瘤细胞存活和增殖;通过MMP-7的活性促进侵袭性和转移过程。通过与表达MUC16的细胞相互作用,也可以促进转移。然而,MSLN与EMT和血管生成相关的基本机制在PDAC中仍然有待阐明。MSLN在80%到90%的PDAC中过度表达,使该靶点成为PDAC患者治疗的一个有吸引力的候选。图2.MSLN在胰腺导管腺癌(PDAC)进展中的作用[2]MSLN与胰腺癌细胞表面的MUC16结合,激活了p38 MAPK依赖性途径,进而上调了MMP-7的合成,导致侵袭和迁移潜力增加。在胰腺癌细胞不表达MUC16的情况下,MSLN能够通过激活ERK依赖性途径上调MMP-7的表达。图3. MSLN-MUC16结合诱导胰腺癌细胞MMP-7的分子通路[3]目前靶向MSLN治疗实体瘤的药物主要有:单克隆抗体药物,携带蛋白毒素的单克隆抗体药物,携带低分子量细胞毒性药物的单克隆抗体药物,靶向MSLN的CAR-T细胞药物,以及可以诱导T细胞针对MSLN产生免疫应答的疫苗。图4. 临床试验中靶向MSLN的方法[4]最新研究进展据不完全统计,目前MSLN靶点处于在研阶段的相关药物有56个,目前进行相关药物研发的企业包括拜耳,Atara Biotherapeutics,诺华在研,亘喜生物,荣昌生物等。目前已有诸多国内外企业在该领域进行了布局。部分处于临床及以上阶段的药物情况统计见下表:数据来源于科睿唯安及公开信息整理针对MSLN靶点机制研究和新药开发的需求,BioMice百奥动物自主研发了B-hMSLN mice和B-hMSLN ID8、B-hMSLN MC38细胞系,助力靶向MSLN药物开发,为临床前药效评估提供了优质模型。B-hMSLN mice基本信息蛋白表达分析利用western blot检测野生型小鼠和B-hMSLN小鼠中种属特异性MSLN的表达。取野生型小鼠(+/+)和杂合B-hMSLN小鼠(H/+)的肺部裂解液,用抗MSLN抗体进行western blot分析。结果显示,小鼠MSLN在杂合B-hMSLN小鼠(H/+)和野生型小鼠(+/+)中均可检测到。人MSLN只在B-hMSLN小鼠(H/+)中可检测到。免疫组化(IHC)方法检测MSLN表达IHC显示在B-hMSLN小鼠肺部组织中有代表性间皮素表达。用人MSLN特异性抗体(A,B)和抗兔IgG抗体(C)特异的抗体对组织进行染色。结果显示,在纯合B-hMSLN小鼠中,细胞膜上显示人MSLN阳性(数据来源于合作方)。B-hMSLN ID8基本信息蛋白表达分析通过流式细胞术对纯合B-hMSLN ID8小鼠细胞中MSLN的表达进行分析。用物种特异性抗MSLN抗体对B-hMSLN ID8培养物的单细胞悬浮液进行染色。在B-hMSLN ID8小鼠细胞的表面检测到人MSLN。B-hMSLN ID8细胞的2-A11克隆被用于体内实验。肿瘤生长曲线&体重变化B-hMSLN ID8小鼠细胞的皮下同种移植肿瘤生长。将B-hMSLN ID8细胞(5x106)和野生型ID8细胞(1x106)皮下植入C57BL/6N小鼠(雌性,6周龄,n=8)。(A)平均肿瘤体积±SEM, (B)体重(平均值±SEM),每周测量两次肿瘤体积和体重,体积以mm3表示,使用公式,V=0.5×长径×短径2。如图A所示,B-hMSLN ID8细胞能够在小鼠体内建立肿瘤,可用于药效研究。肿瘤体积&重量测量肿瘤细胞的蛋白表达分析将B-hMSLN ID8细胞皮下移植到C57BL/6小鼠体内(n=8),在接种后21天,收集肿瘤细胞并通过流式细胞术检测人MSLN的表达。如图所示,人MSLN在肿瘤细胞表面高表达。因此,B-hMSLN ID8小鼠细胞可用于新型MSLN疗法的体内药效研究。B-hMSLN MC38基本信息蛋白表达分析通过流式细胞术对B-hMSLN MC38小鼠进行种属特异性MSLN 表达分析。对 B-hMSLN MC38 培养物的单细胞悬液用种属特异性抗 MSLN 抗体进行染色。结果显示:在B-hMSLN MC38小鼠细胞表面检测到人MSLN,小鼠MSLN不表达。因此 B-hMSLN MC38细胞的1-A03克隆可用于体内实验。肿瘤生长曲线&体重变化B-hMSLN MC38小鼠细胞的皮下同种移植肿瘤生长。B-hMSLN MC38 细胞 (1x106) 和野生型 MC38 细胞 (5x105) 被皮下植入 C57BL/6 小鼠 (雌性, 6 周龄, n=8)。每周两次测量肿瘤体积和体重。(A) 平均肿瘤体积 ± SEM, (B) 体重 (平均值±SEM)。体积以 mm3 表示,使用公式:V=0.5 × 长径 × 短径2。如图 A 所示,B-hMSLN MC38 小鼠细胞能够在体内建立肿瘤,并可用于药效研究。参考文献[1] Mesothelin:An Immunotherapeutic Targetbeyond Solid Tumors. Cancers (Basel). 2022 Mar;14(6):1550.[2] Montemagno C, et al. Int J Mol Sci. 2020 Jun 6;21(11):4067.[3] Chen SH, et al. Sci Rep. 2013;3:1870.[4] Montemagno C, et al. Int J Mol Sci. 2020 Jun 6;21(11):4067.

人类白细胞抗原:HLA,通常称之为MHC分子,是在6号染色体短臂上的一类基因。除同卵双生子以外几乎找不到HLA相同者,每个人的HLA千差万别,是识别人类白细胞的重要的分子标志,是免疫系统区分本身和异体物质的基础,具有非常重要的生物学功能。HLA有不同的基因座,编码两大类MHC蛋白。HLA在医学上的意义主要体现在器官移植,输血,疾病相关性,生理学等方面。人6号染色体HLA基因结构HLA复合体有224个基因座(locus),按其产物的结构、分布与功能分为三群。HLA-I经典I类基因: HLA-A、-B、-C参与递呈内源性抗原。非经典I类基因: HLA-E、-G、-F。HLA-II经典II类基因: HLA-DP 、-DQ、-DR参与递呈外源性抗原。非经典II类基因: LMP、TAP、HLA-DM、HLA-DO 参与抗原的加工和转运。HLA-Ⅲ包括编码补体C4、Bf、C2的基因。编码炎症相关分子TNF、HSP70等基因。HLA基因结构HLA基因功能HLA-I可以提供一般细胞内的一些状况,比如该细胞遭受病毒感染,则将病毒外膜蛋白加工成肽链,通过MHC展示到细胞膜表面,供杀手CD8+ T细胞的识别,以进行扑杀。表达于所有有核细胞表面,例外的仅有神经细胞、胰岛外分泌细胞、心肌细胞和精细胞等。HLA-II可以提供细胞外部的情况,像是组织中有细菌侵入,则巨噬细胞进行吞食后,把细菌蛋白加工成肽链,通过MHC展示给辅助性T细胞,启动体液免疫反应。只位于抗原提呈细胞(APC)表面,例如:B细胞、巨噬细胞、树突状细胞、胸腺上皮细胞和人活化的T细胞等。HLA分子对T细胞在胸腺内的分化成熟过程也起重要作用。体外研究发现:去除胸腺中MHCⅡ类抗原阳性的基质细胞,则CD4+ T细胞的发育受阻,在胸腺培养细胞中加入抗MHCⅡ类抗原的单克隆抗体,也能阻止CD4+ T细胞的发育。目前认为MHC分子在T细胞自身耐受的形成和T细胞库的产生中都起着重要作用。HLA-A2.1分子HLA-A2.1是HLA I 类A基因座,第2复等位基因第1号亚型,备受科学家们的关注。关注HLA-A2.1等位基因的原因有很多,下面将介绍其中的一些原因。T细胞的免疫反应依赖于肽与HLA分子的结合,因此研究者经常研究这些分子与各种抗原的相互作用。由于HLA的高度多态性,有成千上万的HLA分子可供选择,为了获得最大的相关性,研究最常见的表达等位基因是最有意义的。HLA-A2.1的肽结合基序早已为人熟知,HLA多聚体可用于抗原特异性T细胞的染色。甚至有小鼠表达HLA-A2.1等位基因,这使得在更接近人类的生物系统中进行临床前测试成为可能。针对HLA-A2.1分子机制研究的需求,BioMice百奥动物自主研发了B-HLA-A2.1 mice和B-NDG HLA-A2.1 mice,助力HLA-A2.1分子研究,为临床前药效评估提供了优质模型。B-HLA-A2.1 mice基本信息蛋白表达分析流式细胞术检测野生型C57BL/6小鼠(+/+)和纯合型B-HLA-A2.1小鼠(H/H)的脾细胞。野生型C57BL/6小鼠可检测到小鼠B2M和H-2Kb/H-2Db。人B2M和HLA-A2.1只在B-HLA-A2.1纯合小鼠中检测到,而小鼠B2M和H-2Kb/H-2Db在B-HLA-A2.1纯合小鼠中检测不到。 野生C57BL/6 (+/+)小鼠和纯合B-HLA-A2.1小鼠 (H/H)注射抗CD3E抗体后,取脾细胞进行流式细胞术检测。结果显示:野生C57BL/6小鼠可检测到小鼠B2M和H-2Kb/H-2Db。纯合B-HLA-A2.1小鼠中只能检测到人B2M和HLA-A2.1,检测不到小鼠B2M和H-2Kb/H-2Db。脾脏白细胞亚群分析从C57BL/6小鼠和纯合B-HLA-A2.1小鼠(n= 3,8周龄,雌性)中分离脾细胞。流式细胞术分析脾细胞以评估白细胞亚群。结果表明:纯合B-HLA-A2.1小鼠的B细胞、树突状细胞、粒细胞、单核细胞和巨噬细胞的百分率与C57BL/6小鼠相似。CD8+ T细胞百分率显著降低,CD4+ T细胞百分率和NK细胞百分率显著升高,说明hB2M-HLA-A2.1-H-2D替代小鼠B2M可能影响CD8+ T细胞的发育,进而影响脾脏T细胞亚型的比例。数值用平均值±SEM表示。在淋巴结和血液中得到同样的结果。脾脏T细胞亚群分析从C57BL/6和纯合B-HLA-A2.1小鼠(n= 3,8周龄,雌性)中分离脾细胞。流式细胞术分析脾细胞以评估T细胞亚群。结果表明:纯合B-HLA-A2.1小鼠中调节性T细胞的百分率与C57BL/6小鼠相似。CD8+ T细胞百分率显著降低,CD4+ T细胞百分率显著升高,说明hB2M-HLA-A2.1-H-2D替代小鼠B2M可能影响CD8+ T细胞的发育,进而影响脾脏T细胞亚型的比例。数值用平均值±SEM表示。在淋巴结和血液中得到同样的结果。B-NDG HLA-A2.1 mice基本信息蛋白表达分析流式细胞术检测B-NDG小鼠(+/+)和纯合B-NDG HLA-A2.1小鼠(H/H)脾细胞。结果表明:小鼠B2M和H-2Kb/H-2Db在B-NDG小鼠中检测到,但在B-NDG HLA-A2.1小鼠中检测不到。人B2M和HLA-A2.1在纯合B-NDG HLA-A2.1小鼠中检测到,而在B-NDG小鼠中未检测到。脾脏白细胞亚群分析从B-NDG小鼠和B-NDG HLA-A2.1小鼠(n= 3,6周龄,雌性)分离脾细胞。流式细胞术分析脾细胞以评估白细胞亚群。结果表明:纯合B-NDG HLA-A2.1小鼠树突状细胞、粒细胞、单核细胞和巨噬细胞的百分率与B-NDG小鼠相似,表明人源化的B2M和HLA-A2.1不改变这些细胞在脾脏的整体发育、分化和分布。数值用平均值±SEM表示。在骨髓和血液中得到同样的结果。人CD34+HSC免疫重建将人CD34+造血干细胞分别植入经0.8 Gy辐照过的B-NDG HLA-A2.1新生小鼠和B-NDG小鼠。(A) B-NDG HLA-A2.1小鼠存活率略低于B-NDG小鼠,但差异无统计学意义。(B)体重。将人CD34+造血干细胞分别植入经0.8 Gy辐照过的B-NDG HLA-A2.1新生小鼠和B-NDG小鼠。流式细胞术检测人白细胞。结果表明,虽然B-NDG HLA-A2.1小鼠的人CD8+ T细胞比例明显低于B-NDG小鼠,但重建24周内人CD8+ T细胞比例仍维持在18%左右。B-NDG HLA-A2.1小鼠人CD4+ T细胞比例明显高于B-NDG小鼠。B-NDG HLA-A2.1小鼠中其他重组细胞类型的比例与B-NDG小鼠相似。参考资料1、Van Laethem, F., Tikhonova, A.N. & Singer, A. MHC restriction is imposed on a diverse T cell receptor repertoire by CD4 and CD8 co-receptors during thymic selection. Trends in immunology 33, 437-441 (2012).2、Garcia, K.C., Adams, J.J., Feng, D. & Ely, L.K. The molecular basis of TCR germline bias for MHC is surprisingly simple. Nature immunology 10, 143-147 (2009).
肿瘤相关髓系细胞(tumor-associated myeloid cells, TAMC)是肿瘤微环境(TME)中重要的组成部分,具有异质性,在肿瘤微环境中可以发挥不同,甚至是相反的作用,如免疫抑制或免疫刺激。靶向TAMCs作为单一疗法或与化疗、免疫疗法联合应用的研究正在火热进行中。深入研究TAMCs在肿瘤中的功能和作用机制将有助于发现新的治疗方法。研究最多的肿瘤相关髓系细胞包括单核细胞、肿瘤相关巨噬细胞(TAMs)、树突状细胞(DC)、癌症相关循环中性粒细胞、肿瘤相关中性粒细胞(TANs)和骨髓来源抑制细胞(MDSCs)。TAMC的主要功能是调节淋巴细胞行为进而形成免疫刺激或免疫抑制性TME,从而抑制或促进包括肿瘤细胞的恶性克隆进化、生长、存活、侵袭、播散和转移、血管生成在内的各个肿瘤发展阶段。单核细胞单核细胞是一组异质性的单核吞噬细胞,分为经典型单核细胞、中间型单核细胞和非经典型单核细胞,在炎症期间循环外周血中发挥先天免疫功能。肿瘤发展的不同阶段,不同的单核细胞亚群表现出不同甚至相反的作用。从机制上讲,中间型单核细胞经肿瘤细胞刺激后,促炎细胞因子TNF- α和白细胞介素12(IL-12)的产生增加,而抗炎细胞因子白细胞介素10(IL-10)的产生减少,并对肿瘤细胞产生直接的细胞毒性,促进肿瘤细胞凋亡。经典型单核细胞产生VEGF,促进肿瘤细胞外渗,导致转移。相比之下,非经典型单核细胞在吞噬肿瘤细胞衍生微粒后被激活,从而减少肿瘤细胞的转移。肿瘤中单核细胞的功能肿瘤相关巨噬细胞(TAMs)巨噬细胞作为单核吞噬细胞系统,在组织稳态和炎症中起着关键作用。巨噬细胞分为两个主要亚群,M1和M2巨噬细胞,在功能上是异质的。M1巨噬细胞是对抗微生物感染的第一道防线,具有很强的抗原呈递能力诱导强烈的Th1反应。在脂多糖(LPS)、IFN-γ和粒细胞-巨噬细胞刺激因子(GM-CSF)的作用下,M1巨噬细胞经历经典激活,并优先分泌抗菌分子和促炎细胞因子。M2巨噬细胞在限制免疫反应、诱导血管生成和组织修复方面起着关键作用。在白细胞介素4(IL-4)、白细胞介素13(IL- 13)、IL-10和CSF-1的作用下,M2巨噬细胞发生选择性激活并优先分泌抗炎细胞因子。TAMs在肿瘤中的作用树突状细胞(DC)DC是最有效的抗原呈递细胞(APCs),连接先天免疫和适应性免疫,在生理条件下具有表型和功能异质性。在对微生物感染的反应中,细胞外微生物蛋白通常被成熟DC吞噬或内吞,并通过MHC II分子呈递给CD4+ T细胞。相比之下,胞质微生物蛋白通常通过MHC I分子呈现给CD8+ T细胞。TME浸润的DC包括不同发育阶段的树突状细胞亚群,这些肿瘤相关树突状细胞根据不同的细胞亚群和肿瘤分期,发挥免疫刺激或免疫抑制作用。DC在肿瘤中的作用粒细胞粒细胞可大致分为癌症相关循环中性粒细胞、肿瘤相关中性粒细胞(TANs)以及其他中性粒细胞。癌症相关循环中性粒细胞这些与癌症相关的循环中性粒细胞由功能异质性亚群组成,是循环外周血的多形核吞噬细胞,具有对抗微生物病原体的先天免疫功能。在癌症患者中,特别是在晚期和转移后,循环中性粒细胞数量增加,高中性粒细胞与淋巴细胞比率(NLR)与侵袭性癌症相关。肿瘤相关中性粒细胞(TANs)研究表明TME中的TANs可促进肿瘤细胞增殖、外渗和迁移,在肿瘤发展和转移中起着关键作用。TANs可释放弹性蛋白酶等颗粒内容物,促进肿瘤增殖和侵袭细胞;分泌IL-1β和MPPs促进肿瘤细胞外渗到转移前壁龛,从而促进癌细胞的扩散;激活TLR通路,促进肿瘤细胞的迁移、黏附、侵袭和转移。除促肿瘤作用外,TANs还被证明通过产生ROS和TRAIL介导肿瘤细胞的细胞毒性。研究也表明TANs与TME中的淋巴细胞相互作用并调节其功能。其他粒细胞除了嗜中性粒细胞(neutrophilic),其他类型的粒细胞,如嗜酸性粒细胞(eosinophils)和嗜碱性粒细胞(basophils),对肿瘤也有影响。研究表明嗜酸性粒细胞具有抗肿瘤活性,对各种癌细胞都表现出直接的或间接的细胞毒性,从而抑制肿瘤生长。中性粒细胞在肿瘤中的作用骨髓来源抑制细胞(MDSCs)MDSCs是一组异质性的髓系祖细胞和处于不同发育阶段的未成熟髓系细胞,它们在血管中循环。在感染时,MDSCs迅速扩张并分化为粒细胞、单核细胞、巨噬细胞和树突状细胞,在调节免疫反应和组织修复中发挥重要作用。肿瘤相关的MDSC由两个主要亚群体组成,粒细胞骨髓来源抑制细胞(GrMDSCs)和单核细胞骨髓来源抑制细胞(MoMDSCs)。GrMDSCs在表型和形态上与中性粒细胞相似,MoMDSCs在表型和形态上与单核细胞相似,研究表明GrMDSCs和MoMDSCs均参与T细胞介导的免疫抑制。MDSCs及其亚群在肿瘤中的作用髓系细胞在肿瘤免疫治疗的研究中的作用是毋庸置疑的,调节这些髓系细胞的发育、成熟和功能有助于发现新的肿瘤免疫治疗策略。然而,由于各种可互换亚群的复杂性和可塑性,这些髓系细胞执行重叠或相反的功能,目前控制TAMCs行为的分子机制在很大程度上尚不清楚。为进一步阐明髓系细胞各亚群在不同癌症中的功能,并确定其促肿瘤和抗肿瘤活性相关的分子机制,百奥动物自主研发一些列髓系靶点人源化小鼠,助力深入了解髓系细胞的复杂性,并设计新的肿瘤靶向治疗方法。B-hTREM2 mice(C) 基本信息体内药效抗人TREM2抗体在B-hTREM2(C)小鼠中的抗肿瘤活性。纯合子B-hTREM2(C)小鼠(雌性,10周龄,n=6)小鼠皮下接种小鼠乳腺癌EMT-6细胞。当肿瘤体积达到约50-80 mm3时,对小鼠进行分组,然后使用抗人TREM2抗体进行治疗。如图A所示,抗人TREM2抗体在B-hTREM2(C)小鼠中有效地控制肿瘤生长,证实B-hTREM2(C)小鼠模型是抗人TREM2抗体临床前体内药效评估的优质模型。数值以平均值±SEM表示。B-hCD36 mice基本信息体内药效抗小鼠PD-1抗体和抗人CD36抗体联合治疗在B-hCD36小鼠中的抗肿瘤活性。纯合子B-hCD36小鼠(雌性,7周龄,n=5)小鼠皮下接种小鼠结肠癌MC38细胞。当肿瘤体积达到约60 mm3时,对小鼠进行分组,然后使用抗体进行治疗。如图A所示,抗体组合在B-hCD36小鼠中有效地控制肿瘤生长,证实B-hCD36小鼠模式是抗CD36抗体临床前体内药效评估的优质模型。数据来自合作者,数值以平均值±SEM表示。B-hPD-1/hPD-L1/hVSIR mice基本信息蛋白表达分析通过流式细胞术分析野生型C57BL/6小鼠和纯合子B-hPD-1/hPD-L1/hVSIR小鼠中种属特异性PD-1和PD-L1的表达。用抗CD3ε抗体体内刺激野生型C57BL/6小鼠和纯合子B-hPD-1/hPD-L1/hVSIR小鼠,收集脾细胞并进行流式分析。结果显示:小鼠PD-1和PD-L1在野生型C57BL/6小鼠中检测到。人PD-1和PD-L1只在纯合子B-hPD-1/hPD-L1/hVSIR小鼠中检测到,而在野生型C57BL/6小鼠中检测不到。通过流式细胞术分析野生型C57BL/6小鼠和纯合子B-hPD-1/hPD-L1/hVSIR小鼠中种属特异性VSIR的表达。取野生型C57BL/6小鼠和纯合子B-hPD-1/hPD-L1/hVSIR小鼠脾细胞进行流式分析。结果显示:小鼠VSIR仅在野生型C57BL/6小鼠中检测到。人VSIR只在纯合子B-hPD-1/hPD-L1/hVSIR小鼠中检测到,而在野生型C57BL/6小鼠中检测不到。髓系靶点人源化小鼠列表参考资料:1. Aixia Dou, Jing Fan. Heterogeneous Myeloid Cells in Tumors. Cancers (Basel). 2021 Aug; 13(15): 3772.2. Sijin Cheng, Ziyi Li, et al. A pan-cancer single-cell transcriptional atlas of tumor infiltrating myeloid cells. Cell. 2021Feb; 184, 792–809.
康洲大数据 Copyright © 2009-2026 药智网YAOZH.COM All Rights Reserved. 工信部备案号:渝ICP备10200070号-3
客户服务热线:400-678-0778 投诉建议:023-62988285转8001 E-mail:admin@yaozh.com
互联网增值电信业务许可证编号:渝B2-20120028 互联网药品信息服务资格证:(渝)-经营性-2021-0017

渝公网安备 50010802001068号

投诉热线: (023) 6262 8397
邮箱: tousu@yaozh.com
QQ: 236960938

