 药智官方微信
药智官方微信
 药智医械公众号
药智官方微信
药智医械公众号
药智医械公众号
药智官方微信
药智医械公众号

随着生活水平的提高以及医疗技术的不断发展,人类预期寿命在逐步延长,这导致全球老年人口的空前增长,与此同时,与年龄相关的疾病负担也呈指数级增长[1]。有研究指出,在健康状况没有得到改善的情况下单纯延长寿命,是没有意义的[2]。基于此,世界卫生组织在2015年提出了《老龄化与健康全球战略和行动计划》,旨在实现健康老龄化的目标[3]。为了实现这一目标,需要对生物体、器官和细胞水平的衰老及疾病过程有透彻的了解,并针对衰老相关的分子途径开发预防和治疗衰老相关疾病的策略[4]。我国65岁以上人口占总人口比重及预测[5]大多数自发肿瘤、代谢性疾病以及神经退行性疾病的发病率和流行率随年龄增长而增加,其主要原因可以归咎于老年人免疫系统失调。免疫系统可以抵御病原体和杀伤癌细胞。老年人的免疫系统老化,使得免疫反应性急剧下降并且功能失调,这一现象被称为免疫衰老。研究表明,免疫治疗干预在幼小动物身上可能是有效的,但同样的治疗方法在老年动物身上却并不一定有效。考虑到大多数的癌症患者是老年人,癌症的发病率随着老龄化而增加,因此理解免疫系统失调对于免疫治疗的影响,可以更好的干预老年人的癌症治疗进程。为了最优地激发老年人的抗肿瘤免疫反应,需要识别和了解老年人免疫系统的内在缺陷,并使用自身耐受和衰老同时存在的相关模型。[6]小鼠凭借其易于饲养和处理并且寿命较短(~2.5年)的特性,成为衰老及其相关疾病研究中有吸引力的哺乳动物模式生物。[7]百奥动物提供优质老龄鼠(目前最大可提供15月龄,即60周龄的小鼠,陆续可提供最大20-22月龄小鼠),可用于衰老及其相关疾病的研究。可提供品系如下:老龄鼠百奥动物老龄鼠优势① 不同周龄小鼠现货提供;② 高标准的饲养环境;③ 非生产退役种鼠;④ 严格的遗传稳定性监测。C57BL/6J老龄鼠C57BL/6J是使用最为广泛的近交品系之一,也是第一个完成基因组测序,特征分析最全面的小鼠品系。有研究人员对C57BL/6J的生存曲线进行分析,并提供了可比较的对应的人类年龄。衰老是个连续的动态过程,大多数研究将生命阶段简单的分为成年、中年和老年。小鼠月龄和人类年龄对照关系及特点① 小鼠的3-6月龄对应成年人(20-30岁);发育完成尚未受到衰老影响。② 小鼠的10-15月龄对应中年人(38-47岁);可以检测到部分衰老生物标志物(非全部)的衰老变化。③ 小鼠的18-24月龄对应老年人(56-69岁);可以检测到几乎全部衰老标志物的衰老变化[8]。老龄鼠常见表型及病理随着小鼠生长发育,外观会逐渐改变。一些常见的衰老表型有:眼周脱毛、嘴鼻部被理毛、背侧产生脱毛区及灰毛区等。C57BL/6J老龄鼠的部分正常衰老表型[9]同时随着年龄增长,与年龄相关的各项疾病也随之而来,产生一些和疾病相关的表型。例如:自发肿瘤、眼周病变、皮炎脱毛等。非近亲繁殖小鼠长期研究(2年或更长时间)的常见疾病[10]a、Findings from multiple studies are summarized for each line or group. and original terminology.b、4-way crossc、(Af _ C57BL/6)F1d、UM HET3 from (BALB/cByJ _ C57BL/6J)F1 _ (C3H/HeJ _ DBA/2J)F1 老龄鼠应用研究① 健康生命周期的基础生物学研究② 骨质、肌肉、身体组成相关研究③ 饮食、运动、认知与行为研究④ 心血管、内分泌、代谢等疾病研究⑤ 免疫学、肿瘤学、神经退行性疾病研究⑥ 年龄相关的视力、听力障碍研究百奥动物自主研发人源化小鼠模型、疾病模型、老龄鼠等上千种小鼠模型,欢迎来电咨询~参考文献:[1] Olshansky, S.J. From Lifespan to Healthspan. Jama 320, 1323-1324 (2018).[2] Rivera-Tavarez, C.E. Can We Increase Our Health Span? Physical medicine and rehabilitation clinics of North America 28, 681-692 (2017).[3] Beard, J.R. et al. The World report on ageing and health: a policy framework for healthy ageing. Lancet (London, England) 387, 2145-2154 (2016). [4] Barzilai, N., Cuervo, A.M. & Austad, S. Aging as a Biological Target for Prevention and Therapy. Jama 320, 1321-1322 (2018).[5]http://news.youth.cn/jsxw/202103/t20210310_12758403.htm[6] J. Lustgarten.Cancer, aging and immunotherapy: lessons learned from animal models.Cancer Immunol Immunother 58:1979–1989(2009) .[7] Mitchell, S.J., Scheibye-Knudsen, M., Longo, D.L. & de Cabo, R. Animal models of aging research: implications for human aging and age-related diseases. Annual review of animal biosciences 3, 283-303 (2015). [8] Flurkey, K., M. Currer, J. & Harrison, D.E. Chapter 20 - Mouse Models in Aging Research. In: Fox, J.G. et al. (eds). The Mouse in Biomedical Research (Second Edition). Academic Press: Burlington pp 637-672(2007).[9] https:// e. dxy. cn/ broadcast/ live/ id/ 22788/ ? source= wechat& sim= 1& sim_ token= jo3ALEk4m RZ9tfoghOHA rih1tV 6kp0IQvMAdhr7vws7ey9YqHi8uJxdmTk9vElaf[10] C. F. Brayton, P. M. Treuting, and J. M. Ward.Pathobiology of Aging Mice and GEM: Background Strains and Experimental Design.Veterinary Pathology 49(1) 85-105(2012).

1973年,Kobylka和Carraway在乳腺上皮细胞中发现了一种在乳脂球中无法水解的膜蛋白,1978年,该分子被鉴定为血小板GPIV,一种介导血小板中血小板反应蛋白-1(TSP-1)结合的蛋白。随后,Tandon发现GPIV与白细胞分化抗原CD36具有重叠结构。随后,其他研究人员确定了CD36在血小板活化和细胞粘附中的作用。1993年,Abumrad及其同事证明了CD36的脂肪活性,之后将该蛋白在小鼠体内克隆并鉴定为人CD36的小鼠直系同源物,表明其在脂肪酸摄取中的作用。CD36现在被广泛认为是一种清道夫受体,可以吸收长链脂肪酸(LCFAs)和氧化低密度脂蛋白(ox-LDL)。CD36是一种清道夫受体,在多种细胞类型中表达。在脂质代谢中发挥重要作用,介导脂质摄取、免疫识别、炎症、分子粘附和凋亡。与血管生成、炎症反应、动脉粥样硬化性血栓性疾病以及代谢紊乱性糖尿病和肥胖症等有关。在免疫系统中,CD36介导树突状细胞抗原的获取和呈递,并支持调节性T细胞功能。研究表明靶向CD36可能是提高CD8+T细胞免疫疗法的抗肿瘤药效的有效策略,为临床治疗提供了新的思路和可能。[1]CD36与PPAR-β激活[2]CD36靶向作用破坏肿瘤内Treg细胞,并启动PD-1阻断肿瘤。具有抗CD36抗体的MOA: ①耗尽CD36+ Treg细胞。②阻断PPAR-和PPAR-依赖的代谢途径,降低线粒体活性和CD36表达水平,促进Treg细胞凋亡。③阻断TSP-1/CD36通路,促进TME血管生成。CD36调节肿瘤免疫[3]CD36在肿瘤微环境中发挥作用。CD36调节下游Src家族激酶促进抗血管生成、FAO以及化疗耐药和放疗耐药,从而导致肿瘤转移。CD36还通过EMT激活Wnt/TGF-β信号传导促进肿瘤转移。CD36摄取多种脂质,如ox-LDL、LCFAs和胆固醇。免疫细胞中的脂质沉积导致树突状细胞的无菌性炎症和抗原呈递功能障碍,从而诱导肿瘤免疫抑制。此外,CD36可以与凋亡细胞结合并激活交叉引物,这可能导致免疫抑制和肿瘤发展。BioMice百奥动物自主研发了人源化小鼠B-hCD36 mice和人源化细胞系B-hCD36 MC38助力科研。B-hCD36 miceB-hCD36小鼠血常规检测全血细胞计数(CBC)。采集雌性C57BL/6和B-hCD36小鼠(n=8,6-8周龄)的血液并进行全血细胞计数分析。B-hCD36小鼠的测量结果与C57BL/6小鼠相似,表明人源化不会改变血细胞组成和形态。数值表示为平均值±SEM。B-hCD36小鼠血生化检测B-hCD36小鼠的血生化试验。收集C57BL/6和B-hCD36小鼠(n=8,6-8周龄)的血清,并分析其指标水平。B-hCD36小鼠的测量结果与C57BL/6小鼠相似,表明人源化不会改变肝脏等相关组织的健康。数值表示为平均值±SEM。使用抗CD36抗体测试B-hCD36模型的抗体结合试验通过流式细胞术对纯合B-hCD36小鼠进行种属特异性CD36表达分析。收集野生型小鼠 (+/+)和纯合B-hCD36小鼠(H/H)的腹腔渗出巨噬细胞,并通过抗CD36抗体的流式细胞术进行分析。人CD36仅在纯合B-hCD36小鼠中可检测到,但在野生型小鼠中未检测到。CD36-1G04-hIgG1-LALA抗体在小鼠和人中交叉识别。B-hCD36 MC38蛋白表达分析通过流式细胞术对B-hCD36 MC38细胞中的CD36表达进行分析。用种属特异性抗 CD36抗体对B-hCD36 MC38培养物的单细胞混悬液进行染色。在B-hCD36 MC38细胞表面检测到人CD36,但在野生型MC38细胞表面未检测到。使用B-hCD36 MC38细胞的10-G12克隆进行体内实验。肿瘤生长曲线和体重变化B-hCD36 MC38细胞的皮下同种移植肿瘤生长。将B-hCD36 MC38细胞(5x105)和野生型MC38细胞(5x105)皮下植入B-hCD36小鼠(雌性,7周龄,n=5)。每周测量两次肿瘤体积和体重。(A)平均肿瘤体积±SEM。(B)体重(平均值±SEM)。体积以mm3表示,使用公式:V=0.5X长径X短径2。如图A所示,B-hCD36 MC38细胞能够在体内建立肿瘤,可用于有效性研究。肿瘤细胞蛋白表达分析将B-hCD36 MC38细胞皮下移植到B-hCD36小鼠(n=5)中,接种后35天,收获肿瘤细胞并通过流式细胞术评估人CD36表达。如图所示,人CD36在肿瘤细胞表面高度表达。因此,B-hCD36 MC38细胞可用于新型CD36疗法的体内药效研究。更多验证数据信息,正在研究进行中,敬请关注。想要获取更多百奥动物自主研发的人源化小鼠信息,请访问百奥动物官网或扫描下方二维码。参考文献1.CD36-mediated ferroptosis dampens intratumoral CD8+T cell effector function and impairs their antitumor ability2.Wang, H., Franco, F., Tsui, Y. et al. CD36-mediated metabolic adaptation supports regulatory T cell survival and function in tumors. Nat Immunol 21, 298–308 (2020). doi:10.1038/s41590-019-0589-53.Wang, Jingchun; Li, Yongsheng (2019). CD36 tango in cancer: signaling pathways and functions. Theranostics, 9(17), 4893–4908. doi:10.7150/thno.36037
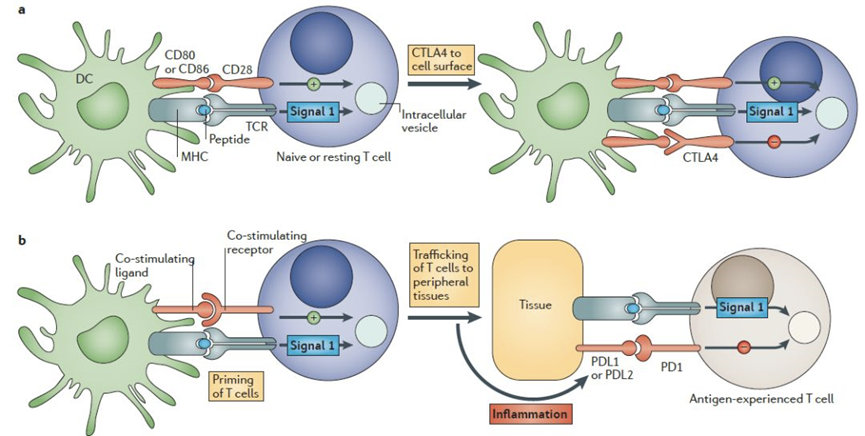
2006年3月,TeGenero公司的CD28超级激动剂单抗TGN1412治疗风湿性关节炎/白血病的1期临床试验中,6名志愿者遭受细胞因子释放综合征并伴有多器官功能衰竭,全身极度肿胀成为“大象人”,万幸的是最终脱离生命危险,该事件直接导致TeGenero破产。16年前的黑天鹅事件也给CD28药物开发蒙上了一层阴影。后续TGN1412并未被就此放弃,莫斯科抗体公司TheraMab顺利接盘并更名为TAB08,有报道称1期临床NCT03006029、NCT01990157不良事件保持在一个可接受的水平(短暂的发热与IL-6有关),然而试验如今已经因“行政原因”终止,这款CD28单抗最终也没能迎来涅槃重生[1]。如此看来,再次沉沦的CD28的未来当真就是砂砾宿命了吗?2017年,Science上发表了一篇题为Rescue of exhausted CD8 Tcells by PD-1 targeted therapies is CD28-dependent的文章,证明了CD28/B7共刺激途径对有效的PD-1疗法至关重要。至此,CD28再次回到研究者的视线中。2019年11月,赛诺菲在Nature Cancer发表了一款CD28/CD3/CD38三抗的研究进展。该三抗在CD3分子亲和力的选择上,采用了中等亲和力的抗体(KD~20nM)平衡了杀伤有效性以及细胞因子大量释放的安全性问题[2]。2020年1月,再生元在Science Translational Medicine发表论文,指出CD28双抗可以增强CD3的抗肿瘤疗效。在动物实验中,共刺激型CD28双抗明显增强了CD3双抗的疗效,且没有细胞因子风暴的风险。2020年6月24日,再生元再次在ScienceTranslational Medicine杂志上发表论文,证实了肿瘤特异性抗原(TSA) x CD28双特异性抗体可以与更广泛的抗PD-1抗体协同作用,增强癌症治疗效果,诱导长效抗肿瘤免疫力,且不会诱发细胞因子风暴,具有很好的耐受性。今年2月,赛诺菲的研发团队在Nature发表了一篇HER2×CD3×CD28三抗的研究进展。研究表明,在原代人CD3+T细胞重组的免疫缺陷NSG小鼠体内,该三抗使CD8 T细胞中颗粒酶的表达增加了6.8倍。相关肿瘤药效实验结果表明,剂量低至10μg/kg时,HER2×CD3×CD28三抗依然能够诱导肿瘤消退。近期,惠和生物靶向CD3×CD28×CD19三特异性抗体CC312的IND申请获得美国FDA默示许可,进入临床研发阶段,CC312是国内首个,也是全球第三个基于CD28共刺激信号的三特异性抗体。随着赛诺菲、再生元等牵头的研究推进,CD28被强势拉回公众视野,更多的潜力有待开发!CD28相关信号通路CD28是T细胞激活最重要的参与者之一,是增强MHC-TCR的主激活信号的主要辅助信号。CD28和CTLA-4具有高度的同源性,具有相同的配体CD80和CD86(B7-1和B7-2),CD28负责传递激活信号,活化T细胞,而CTLA-4负责传递抑制信号给T细胞,让T细胞不会杀伤其它细胞,包括肿瘤细胞。CTLA-4与配体的亲和力要高于CD28,因此会竞争阻断CD28的T细胞激活作用。在T细胞辅助激活的过程中,CD86优先表达,在与CD28结合上,CD86表现出优于CD80的结合能力,有利于T细胞激活。在T细胞激活的过程中,CD28扮演着“加油”角色,CTLA-4扮演着“前刹车”角色,PD-1则扮演着“后刹车”角色[3]。CD28、CTLA-4、PD-1/PD-L1信号通路[4]CD28靶点部分开发进展CD28作为一个老靶点,不仅机制清晰,而且针对不同临床适应症,既可开发拮抗剂也可开发激动剂,理应前景无限,但由于其研发历程的跌宕起伏,目前只有少量抗体药物开发管线。其临床药物开发上主要集中在3个方面:CD28关联靶点融合蛋白药物;单抗药物;多特异性抗体药物。(数据来源科睿唯安及公开信息整理)CD28系列人源化动物模型对于CD28相关药物的研究,再生元和赛诺菲已经做出了很好的表率,进一步的数据也许会让CD28再次火热起来。相关动物模型对于CD28靶向调节剂开发可谓至关重要,BioMice 百奥动物自主研发的CD28系列人源化鼠是评估CD28相关抗体药物的优质临床前实验动物模型。B-hCD28 小鼠验证数据蛋白表达分析流式细胞术分析B-hCD28纯合小鼠CD28的表达取抗CD3ε抗体刺激野生型小鼠和纯合B-hCD28小鼠的脾细胞,用种属特异性抗CD28抗体进行流式分析。小鼠CD28在野生型小鼠中检测到。人CD28仅在纯合B-hCD28小鼠中检测到,而在野生型小鼠中检测不到。抗人CD28抗体的体内药效抗人CD28抗体在B-hCD28小鼠体内的抗肿瘤活性实验结果表明,两种抗人CD28抗体在B-hCD28小鼠体内均能有效控制肿瘤生长,表明B-hCD28小鼠为抗人CD28抗体的体内评价提供了一个优质的临床前模型。抗人PD-L1xCD28双特异性抗体的体内药效抗PD-L1和抗CD28 (PD-L1xCD28)双特异性抗体(BsAb)在B-hCD28小鼠中的抗肿瘤活性结果表明,抗人PD-L1xCD28 BsAbs能够有效控制B-hCD28小鼠的肿瘤生长,表明B-hCD28小鼠为抗人PD-L1xCD28 BsAbs的体内评价提供了一个优质的临床前模型。B-hCD3E/hCD28小鼠验证数据蛋白表达分析流式细胞术分析B-hCD3E/hCD28纯合小鼠CD3E的表达取野生型小鼠和纯合B-hCD3E/hCD28小鼠脾细胞,用种属特异性抗CD3E抗体进行流式细胞术分析。小鼠CD3E在野生型小鼠中检测到。人CD3E仅在纯合B-hCD3E/hCD28小鼠中检测到,而在野生型小鼠中检测不到。流式细胞术分析B-hCD3E/hCD28纯合小鼠CD28的表达取野生型小鼠和纯合B-hCD3E/hCD28小鼠脾细胞,用种属特异性抗CD28抗体进行流式细胞术分析。小鼠CD28在野生型小鼠中检测到。人CD28仅在纯合B-hCD3E/hCD28小鼠中检测到,而在野生型小鼠中检测不到。B-hCD3EDG/hCD28 小鼠验证数据蛋白表达分析流式细胞术分析B-hCD3EDG/hCD28纯合小鼠CD3E的表达取野生型小鼠和纯合B-hCD3EDG/hCD28小鼠脾细胞,用种属特异性抗CD3E抗体进行流式分析。小鼠CD3E在野生型小鼠中检测到。人CD3E仅在纯合B-hCD3EDG/hCD28小鼠中检测到,而在野生型小鼠中检测不到。流式细胞术分析B-hCD3EDG/hCD28纯合小鼠CD28的表达取野生型小鼠和纯合B-hCD3EDG/hCD28小鼠脾细胞,用种属特异性抗CD28抗体进行流式细胞术分析。小鼠CD28在野生型小鼠中检测到。人CD28仅在纯合B-hCD3EDG/hCD28小鼠中检测到,而在野生型小鼠中检测不到。B-hCD28/hTROP2小鼠验证数据蛋白表达分析流式细胞术分析B-hCD28/hTROP2纯合小鼠CD28的表达取野生型小鼠和纯合B-hCD28/hTROP2小鼠脾细胞,用种属特异性抗CD28抗体进行流式细胞术分析。小鼠CD28在野生型小鼠中检测到。人CD28仅在纯合B-hCD28/hTROP2小鼠中检测到,而在野生型小鼠中检测不到。Western blot分析TROP2在B-hCD28/hTROP2纯合小鼠中的表达取野生型小鼠和纯合B-hCD28/hTROP2小鼠的皮肤组织,用抗TROP2抗体进行western blot分析。小鼠TROP2在野生型小鼠中检测到。人TROP2仅在纯合B-hCD28/hTROP2小鼠中检测到,而在野生型小鼠中检测不到。B-hCD28/hB7-H3小鼠验证数据蛋白表达分析流式细胞术分析B-hCD28/hB7-H3纯合小鼠CD28的表达取野生型小鼠和纯合B-hCD28/hB7-H3小鼠脾细胞,用种属特异性抗CD28抗体进行流式细胞术分析。小鼠CD28在野生型小鼠中检测到。人CD28仅在纯合B-hCD28/hB7-H3小鼠中检测到,而在野生型小鼠中检测不到。Western blot分析B-hCD28/hB7-H3纯合小鼠B7-H3的表达采集野生型小鼠和纯合B-hCD28/hB7-H3小鼠附睾,用抗B7-H3抗体进行western blot分析。由于抗体的交叉反应,野生型小鼠和纯合B-hCD28/hB7-H3小鼠均可检测到B7-H3。B-hCD28/hCD20小鼠验证数据蛋白表达分析流式细胞术分析B-hCD28/hCD20纯合小鼠CD28的表达取野生型小鼠和纯合B-hCD28/hCD20小鼠脾细胞,用种属特异性抗CD28抗体进行流式细胞术分析。小鼠CD28在野生型小鼠中检测到。人CD28仅在纯合B-hCD28/hCD20小鼠中检测到,而在野生型小鼠中检测不到。流式细胞术检测B-hCD28/hCD20纯合小鼠CD20的表达取野生型小鼠和纯合B-hCD28/hCD20 小鼠脾细胞,用种属特异性抗CD20抗体进行流式细胞术分析。小鼠CD20在野生型小鼠中检测到。人CD20仅在纯合B-hCD28/hCD20小鼠中检测到,而在野生型小鼠中检测不到。B-hSIRPA/hCD47/hCD3E/hCD28小鼠验证数据蛋白表达分析流式细胞术分析B-hSIRPA/hCD47/hCD3E/hCD28纯合小鼠CD3E、CD28、SIRPA、CD47的表达取野生型小鼠和纯合B-hSIRPA/hCD47/hCD3E/hCD28小鼠脾细胞,分别用种特异性抗CD3E、CD28、SIRPA、CD47抗体进行流式细胞术分析。由于抗体的交叉反应,野生型小鼠和B-hSIRPA/hCD47/hCD3E/hCD28小鼠均可检测到小鼠SIRPA,人SIRPA仅在纯合B-hSIRPA/hCD47/hCD3E/hCD28小鼠中检测到,而在野生型小鼠中未检测到;小鼠CD3E、CD28、CD47在野生型小鼠中检测到,人CD3E、CD28、CD47仅在纯合B-hSIRPA/hCD47/hCD3E/hCD28小鼠中检测到,而在野生型小鼠中检测不到。CD28靶点相关模型列表 想要了解更多人源化小鼠,欢迎扫描下方二维码查看或者来电咨询。 参考资料:[1] https://mp.weixin.qq.com/s/6tuPYtTIr_gNQrb0N7o7_w[2] Sanofi R&D Investor Event[3] Dimitris Skokos et al. A class of costimulatoryCD28-bispecific antibodies that enhance the antitumor activity ofCD3-bispecific antibodies. Sci. Transl. Med. 2020[4]https://oncologypro.esmo.org/education-library/esmo-handbooks/immuno-oncology/immune-synapse
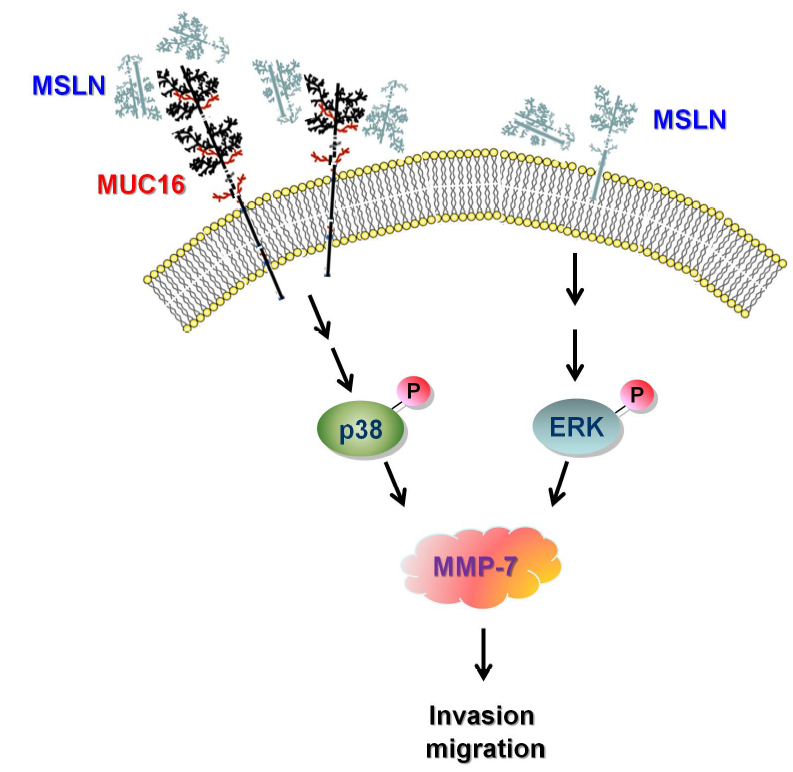
MSLN基因编码一种前体蛋白,经蛋白水解处理生成两种蛋白产物,主要位于正常间皮细胞表面,两种蛋白产物为巨核细胞增强因子(megakaryocyte potentiating,MPF)和间皮素(Mesothelin,MSLN)。巨核细胞增强因子作为细胞因子可刺激骨髓巨核细胞集落形成。间皮素在正常组织中,仅表达在间皮细胞,但在间皮瘤(mesothelioma)、卵巢癌(ovarian cancer)、胰腺癌(pancreatic cancer)、胆管癌(cholangiocarcinoma)等肿瘤中均有高表达。因此间皮素是很有前途的肿瘤特异性治疗候选药物靶点。图1.间皮素的结构和功能[1]MSLN通过ERK和PI3K/Akt通路促进肿瘤细胞存活和增殖;通过MMP-7的活性促进侵袭性和转移过程。通过与表达MUC16的细胞相互作用,也可以促进转移。然而,MSLN与EMT和血管生成相关的基本机制在PDAC中仍然有待阐明。MSLN在80%到90%的PDAC中过度表达,使该靶点成为PDAC患者治疗的一个有吸引力的候选。图2.MSLN在胰腺导管腺癌(PDAC)进展中的作用[2]MSLN与胰腺癌细胞表面的MUC16结合,激活了p38 MAPK依赖性途径,进而上调了MMP-7的合成,导致侵袭和迁移潜力增加。在胰腺癌细胞不表达MUC16的情况下,MSLN能够通过激活ERK依赖性途径上调MMP-7的表达。图3. MSLN-MUC16结合诱导胰腺癌细胞MMP-7的分子通路[3]目前靶向MSLN治疗实体瘤的药物主要有:单克隆抗体药物,携带蛋白毒素的单克隆抗体药物,携带低分子量细胞毒性药物的单克隆抗体药物,靶向MSLN的CAR-T细胞药物,以及可以诱导T细胞针对MSLN产生免疫应答的疫苗。图4. 临床试验中靶向MSLN的方法[4]最新研究进展据不完全统计,目前MSLN靶点处于在研阶段的相关药物有56个,目前进行相关药物研发的企业包括拜耳,Atara Biotherapeutics,诺华在研,亘喜生物,荣昌生物等。目前已有诸多国内外企业在该领域进行了布局。部分处于临床及以上阶段的药物情况统计见下表:数据来源于科睿唯安及公开信息整理针对MSLN靶点机制研究和新药开发的需求,BioMice百奥动物自主研发了B-hMSLN mice和B-hMSLN ID8、B-hMSLN MC38细胞系,助力靶向MSLN药物开发,为临床前药效评估提供了优质模型。B-hMSLN mice基本信息蛋白表达分析利用western blot检测野生型小鼠和B-hMSLN小鼠中种属特异性MSLN的表达。取野生型小鼠(+/+)和杂合B-hMSLN小鼠(H/+)的肺部裂解液,用抗MSLN抗体进行western blot分析。结果显示,小鼠MSLN在杂合B-hMSLN小鼠(H/+)和野生型小鼠(+/+)中均可检测到。人MSLN只在B-hMSLN小鼠(H/+)中可检测到。免疫组化(IHC)方法检测MSLN表达IHC显示在B-hMSLN小鼠肺部组织中有代表性间皮素表达。用人MSLN特异性抗体(A,B)和抗兔IgG抗体(C)特异的抗体对组织进行染色。结果显示,在纯合B-hMSLN小鼠中,细胞膜上显示人MSLN阳性(数据来源于合作方)。B-hMSLN ID8基本信息蛋白表达分析通过流式细胞术对纯合B-hMSLN ID8小鼠细胞中MSLN的表达进行分析。用物种特异性抗MSLN抗体对B-hMSLN ID8培养物的单细胞悬浮液进行染色。在B-hMSLN ID8小鼠细胞的表面检测到人MSLN。B-hMSLN ID8细胞的2-A11克隆被用于体内实验。肿瘤生长曲线&体重变化B-hMSLN ID8小鼠细胞的皮下同种移植肿瘤生长。将B-hMSLN ID8细胞(5x106)和野生型ID8细胞(1x106)皮下植入C57BL/6N小鼠(雌性,6周龄,n=8)。(A)平均肿瘤体积±SEM, (B)体重(平均值±SEM),每周测量两次肿瘤体积和体重,体积以mm3表示,使用公式,V=0.5×长径×短径2。如图A所示,B-hMSLN ID8细胞能够在小鼠体内建立肿瘤,可用于药效研究。肿瘤体积&重量测量肿瘤细胞的蛋白表达分析将B-hMSLN ID8细胞皮下移植到C57BL/6小鼠体内(n=8),在接种后21天,收集肿瘤细胞并通过流式细胞术检测人MSLN的表达。如图所示,人MSLN在肿瘤细胞表面高表达。因此,B-hMSLN ID8小鼠细胞可用于新型MSLN疗法的体内药效研究。B-hMSLN MC38基本信息蛋白表达分析通过流式细胞术对B-hMSLN MC38小鼠进行种属特异性MSLN 表达分析。对 B-hMSLN MC38 培养物的单细胞悬液用种属特异性抗 MSLN 抗体进行染色。结果显示:在B-hMSLN MC38小鼠细胞表面检测到人MSLN,小鼠MSLN不表达。因此 B-hMSLN MC38细胞的1-A03克隆可用于体内实验。肿瘤生长曲线&体重变化B-hMSLN MC38小鼠细胞的皮下同种移植肿瘤生长。B-hMSLN MC38 细胞 (1x106) 和野生型 MC38 细胞 (5x105) 被皮下植入 C57BL/6 小鼠 (雌性, 6 周龄, n=8)。每周两次测量肿瘤体积和体重。(A) 平均肿瘤体积 ± SEM, (B) 体重 (平均值±SEM)。体积以 mm3 表示,使用公式:V=0.5 × 长径 × 短径2。如图 A 所示,B-hMSLN MC38 小鼠细胞能够在体内建立肿瘤,并可用于药效研究。参考文献[1] Mesothelin:An Immunotherapeutic Targetbeyond Solid Tumors. Cancers (Basel). 2022 Mar;14(6):1550.[2] Montemagno C, et al. Int J Mol Sci. 2020 Jun 6;21(11):4067.[3] Chen SH, et al. Sci Rep. 2013;3:1870.[4] Montemagno C, et al. Int J Mol Sci. 2020 Jun 6;21(11):4067.

人类白细胞抗原:HLA,通常称之为MHC分子,是在6号染色体短臂上的一类基因。除同卵双生子以外几乎找不到HLA相同者,每个人的HLA千差万别,是识别人类白细胞的重要的分子标志,是免疫系统区分本身和异体物质的基础,具有非常重要的生物学功能。HLA有不同的基因座,编码两大类MHC蛋白。HLA在医学上的意义主要体现在器官移植,输血,疾病相关性,生理学等方面。人6号染色体HLA基因结构HLA复合体有224个基因座(locus),按其产物的结构、分布与功能分为三群。HLA-I经典I类基因: HLA-A、-B、-C参与递呈内源性抗原。非经典I类基因: HLA-E、-G、-F。HLA-II经典II类基因: HLA-DP 、-DQ、-DR参与递呈外源性抗原。非经典II类基因: LMP、TAP、HLA-DM、HLA-DO 参与抗原的加工和转运。HLA-Ⅲ包括编码补体C4、Bf、C2的基因。编码炎症相关分子TNF、HSP70等基因。HLA基因结构HLA基因功能HLA-I可以提供一般细胞内的一些状况,比如该细胞遭受病毒感染,则将病毒外膜蛋白加工成肽链,通过MHC展示到细胞膜表面,供杀手CD8+ T细胞的识别,以进行扑杀。表达于所有有核细胞表面,例外的仅有神经细胞、胰岛外分泌细胞、心肌细胞和精细胞等。HLA-II可以提供细胞外部的情况,像是组织中有细菌侵入,则巨噬细胞进行吞食后,把细菌蛋白加工成肽链,通过MHC展示给辅助性T细胞,启动体液免疫反应。只位于抗原提呈细胞(APC)表面,例如:B细胞、巨噬细胞、树突状细胞、胸腺上皮细胞和人活化的T细胞等。HLA分子对T细胞在胸腺内的分化成熟过程也起重要作用。体外研究发现:去除胸腺中MHCⅡ类抗原阳性的基质细胞,则CD4+ T细胞的发育受阻,在胸腺培养细胞中加入抗MHCⅡ类抗原的单克隆抗体,也能阻止CD4+ T细胞的发育。目前认为MHC分子在T细胞自身耐受的形成和T细胞库的产生中都起着重要作用。HLA-A2.1分子HLA-A2.1是HLA I 类A基因座,第2复等位基因第1号亚型,备受科学家们的关注。关注HLA-A2.1等位基因的原因有很多,下面将介绍其中的一些原因。T细胞的免疫反应依赖于肽与HLA分子的结合,因此研究者经常研究这些分子与各种抗原的相互作用。由于HLA的高度多态性,有成千上万的HLA分子可供选择,为了获得最大的相关性,研究最常见的表达等位基因是最有意义的。HLA-A2.1的肽结合基序早已为人熟知,HLA多聚体可用于抗原特异性T细胞的染色。甚至有小鼠表达HLA-A2.1等位基因,这使得在更接近人类的生物系统中进行临床前测试成为可能。针对HLA-A2.1分子机制研究的需求,BioMice百奥动物自主研发了B-HLA-A2.1 mice和B-NDG HLA-A2.1 mice,助力HLA-A2.1分子研究,为临床前药效评估提供了优质模型。B-HLA-A2.1 mice基本信息蛋白表达分析流式细胞术检测野生型C57BL/6小鼠(+/+)和纯合型B-HLA-A2.1小鼠(H/H)的脾细胞。野生型C57BL/6小鼠可检测到小鼠B2M和H-2Kb/H-2Db。人B2M和HLA-A2.1只在B-HLA-A2.1纯合小鼠中检测到,而小鼠B2M和H-2Kb/H-2Db在B-HLA-A2.1纯合小鼠中检测不到。 野生C57BL/6 (+/+)小鼠和纯合B-HLA-A2.1小鼠 (H/H)注射抗CD3E抗体后,取脾细胞进行流式细胞术检测。结果显示:野生C57BL/6小鼠可检测到小鼠B2M和H-2Kb/H-2Db。纯合B-HLA-A2.1小鼠中只能检测到人B2M和HLA-A2.1,检测不到小鼠B2M和H-2Kb/H-2Db。脾脏白细胞亚群分析从C57BL/6小鼠和纯合B-HLA-A2.1小鼠(n= 3,8周龄,雌性)中分离脾细胞。流式细胞术分析脾细胞以评估白细胞亚群。结果表明:纯合B-HLA-A2.1小鼠的B细胞、树突状细胞、粒细胞、单核细胞和巨噬细胞的百分率与C57BL/6小鼠相似。CD8+ T细胞百分率显著降低,CD4+ T细胞百分率和NK细胞百分率显著升高,说明hB2M-HLA-A2.1-H-2D替代小鼠B2M可能影响CD8+ T细胞的发育,进而影响脾脏T细胞亚型的比例。数值用平均值±SEM表示。在淋巴结和血液中得到同样的结果。脾脏T细胞亚群分析从C57BL/6和纯合B-HLA-A2.1小鼠(n= 3,8周龄,雌性)中分离脾细胞。流式细胞术分析脾细胞以评估T细胞亚群。结果表明:纯合B-HLA-A2.1小鼠中调节性T细胞的百分率与C57BL/6小鼠相似。CD8+ T细胞百分率显著降低,CD4+ T细胞百分率显著升高,说明hB2M-HLA-A2.1-H-2D替代小鼠B2M可能影响CD8+ T细胞的发育,进而影响脾脏T细胞亚型的比例。数值用平均值±SEM表示。在淋巴结和血液中得到同样的结果。B-NDG HLA-A2.1 mice基本信息蛋白表达分析流式细胞术检测B-NDG小鼠(+/+)和纯合B-NDG HLA-A2.1小鼠(H/H)脾细胞。结果表明:小鼠B2M和H-2Kb/H-2Db在B-NDG小鼠中检测到,但在B-NDG HLA-A2.1小鼠中检测不到。人B2M和HLA-A2.1在纯合B-NDG HLA-A2.1小鼠中检测到,而在B-NDG小鼠中未检测到。脾脏白细胞亚群分析从B-NDG小鼠和B-NDG HLA-A2.1小鼠(n= 3,6周龄,雌性)分离脾细胞。流式细胞术分析脾细胞以评估白细胞亚群。结果表明:纯合B-NDG HLA-A2.1小鼠树突状细胞、粒细胞、单核细胞和巨噬细胞的百分率与B-NDG小鼠相似,表明人源化的B2M和HLA-A2.1不改变这些细胞在脾脏的整体发育、分化和分布。数值用平均值±SEM表示。在骨髓和血液中得到同样的结果。人CD34+HSC免疫重建将人CD34+造血干细胞分别植入经0.8 Gy辐照过的B-NDG HLA-A2.1新生小鼠和B-NDG小鼠。(A) B-NDG HLA-A2.1小鼠存活率略低于B-NDG小鼠,但差异无统计学意义。(B)体重。将人CD34+造血干细胞分别植入经0.8 Gy辐照过的B-NDG HLA-A2.1新生小鼠和B-NDG小鼠。流式细胞术检测人白细胞。结果表明,虽然B-NDG HLA-A2.1小鼠的人CD8+ T细胞比例明显低于B-NDG小鼠,但重建24周内人CD8+ T细胞比例仍维持在18%左右。B-NDG HLA-A2.1小鼠人CD4+ T细胞比例明显高于B-NDG小鼠。B-NDG HLA-A2.1小鼠中其他重组细胞类型的比例与B-NDG小鼠相似。参考资料1、Van Laethem, F., Tikhonova, A.N. & Singer, A. MHC restriction is imposed on a diverse T cell receptor repertoire by CD4 and CD8 co-receptors during thymic selection. Trends in immunology 33, 437-441 (2012).2、Garcia, K.C., Adams, J.J., Feng, D. & Ely, L.K. The molecular basis of TCR germline bias for MHC is surprisingly simple. Nature immunology 10, 143-147 (2009).
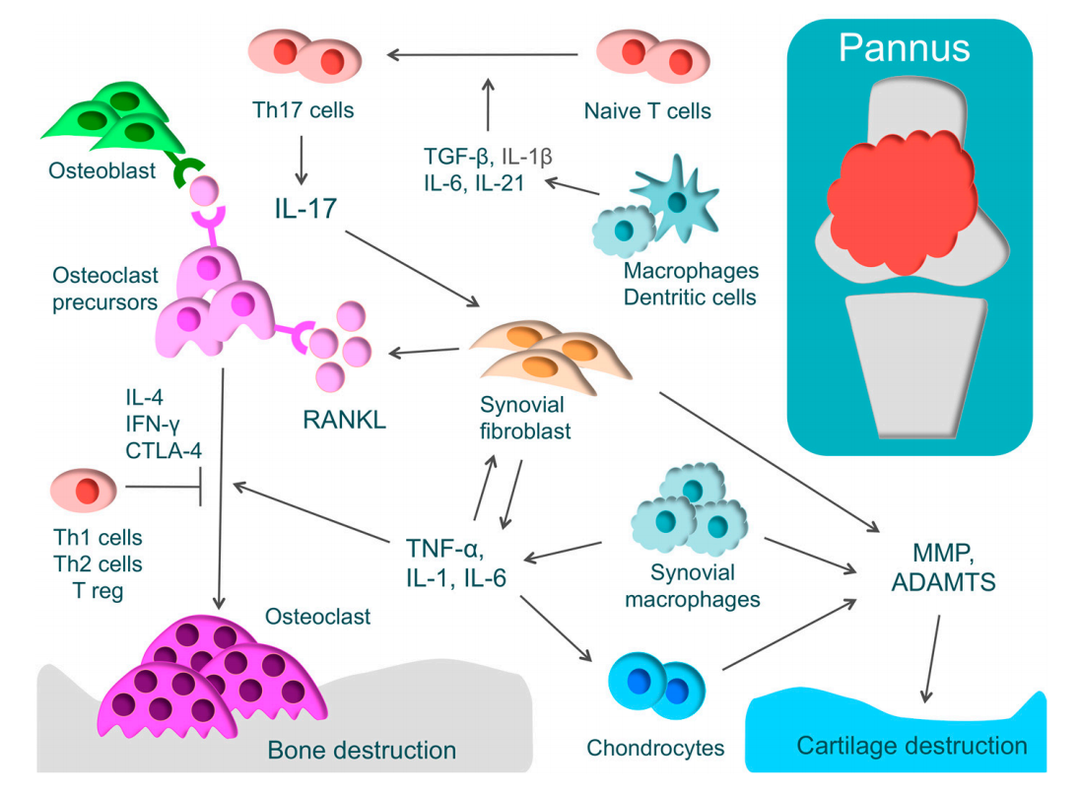
不死的癌症类风湿性关节炎(Rheumatoid Arthritis,RA)是人类最常见的自身免疫性疾病之一,其临床症状主要表现有全身关节肿胀、疼痛和运动失调等,病情极其严重的甚至可能会导致残疾。而病理主要表现出增生性滑膜炎,软骨损伤以及骨结构的破坏。为了更好的研究人类的类风湿性关节炎,研究者在小鼠上开发出多种能模拟人类类风湿性关节炎的动物模型,而这之中胶原诱导型关节炎,能最大程度反映出人类类风湿性关节炎的临床和病理特征。Rheumatoid Arthritis百奥动物在不同品系的小鼠(C57BL/6、DBA)上都建立了稳定的胶原诱导型关节炎疾病模型(Collagen-Induced Arthritis,CIA),可用于相关药物的药效评价。类风湿性关节炎 (RA) 的病理及软骨和骨破坏的机制RA 的特征是增生性滑膜(血管翳)和 T 细胞的过度免疫反应。血管翳包括 T 细胞、滑膜成纤维细胞和巨噬细胞,可产生炎性细胞因子,如肿瘤坏死因子 (TNF)-α、白细胞介素-1(IL-1)、IL-6和IL-17。相关数据展示01RA/CIA 模型介绍 临床评分利用胶原(CII)在C57BL/6小鼠中诱导关节炎模型。(A)小鼠体重变化;(B)临床评分;(C)小鼠发病率。结果显示:在造模组(G2)中,小鼠的临床评分显著增加,提示关节炎模型建立成功。 病理分析C57BL/6小鼠关节炎模型建立后的病理分析。(A)病理评分;(B)病理切片H&E染色。造模组动物全部或部分关节可见皮下混合炎细胞浸润(a),关节周围狭窄(b),踝关节和/或趾骨关节软骨和骨组织破坏(c)等关节炎病变,进一步提示关节炎模型建立成功。02C57BL/6小鼠关节炎模型用于评价甲氨蝶呤(MTX)药效 临床评分甲氨蝶呤在小鼠关节炎模型中的药效作用。(A)小鼠体重变化;(B)临床评分;(C)小鼠发病率。结果显示:在造模组(G2)中,小鼠的临床评分显著增加,表明CIA模型建立成功,同时相比于未造模组(G1),体重波动更加明显。在给于甲氨喋呤(MTX)后,治疗组(G3)的平均临床评分显著低于造模组,证明小鼠的病情得到了有效控制。另外,在发病率方面,治疗组的发病率最大值为40%,也明显低于造模组近80%的发病率,提示小分子药甲氨喋呤对该疾病具有治疗作用。 病理分析甲氨喋呤在小鼠关节炎模型中药效作用的病理分析。(A)病理评分;(B)病理切片H&E染色。病理结果显示:未造模组(G1)动物镜下未见明显异常改变,踝关节软骨表面光滑,关节腔明显(a)。造模组(G2)动物踝关节周围组织,可见皮下混合炎细胞浸润,关节滑膜炎和/或血管翳形成(c),关节软骨破坏,关节腔消失,部分骨组织融合(b)。与G2造模组相比,治疗组虽然有部分踝关节周围皮下组织水肿和炎细胞浸润(d),但其病理评分均值明显低于G2组,甲氨喋呤对动物关节炎病变具有治疗作用。03B-hIL6/hIL6R小鼠关节炎模型用于评价抗人IL6抗体sirukumab药效 临床评分抗人IL-6抗体sirukumab(内部合成)在小鼠关节炎模型中药效作用。(A)小鼠体重变化;(B)临床评分。结果显示:在模型成功建立后,治疗组(G3,G4)小鼠用sirukumab抗体进行治疗,显示出治疗作用,且表现出剂量依赖性。 病理分析抗人IL-6抗体sirukumab(内部合成)在小鼠关节炎模型中药效作用的病理分析。(A)病理切片H&E染色;(B)病理评分。结果显示:未造模组(G1)动物未见明显异常改变,踝关节软骨表面光滑,关节腔明显。造模组(G2)动物踝关节骨组织损伤(d),关节腔或关节周围腔隙消失(e),并出现血管翳(a)。与造模组相比,抗体药治疗低剂量组(G3)有部分炎细胞浸润(b)和血管翳(a)生成,并表现出滑膜增生(c)。但在抗体药治疗高剂量组(G4),仅有部分血管翳,关节炎病变基本消失,关节腔明显,提示sirukumab对小鼠关节炎具有治疗作用。04服务项目更多药效相关服务,CAIA引起的关节炎模型,骨关节炎模型,骨质疏松模型,正在研究进行中,感兴趣的欢迎联系我们获取资料。想要获取更多百奥动物自主研发的自身免疫性疾病模型信息,请访问百奥动物官网。参考文献:1. Tateiwa, D., Yoshikawa, H. & Kaito, T. Cartilage and Bone Destruction in Arthritis: Pathogenesis and Treatment Strategy: A Literature Review. Cells 8, doi:10.3390/cells8080818 (2019).2. McInnes IB, et al. Cytokines in the pathogenesis of rheumatoid arthritis. Nat Rev Immunol 7, 429-42 (2007).
医药生物行业生命科学领域产业链系列报告显示,随着科研发展与对新靶点的持续追求,基因修饰动物凭借可以精准满足用户需求的特点,正快速发展为模式动物市场主流产品。GMI 数据披露,2019 年全球基因修饰动物模型市场规模约为 100 亿美元,预计 2023 年将增至 141 亿美元,占动物模型服务市场近67%[1]。技术进步重塑市场格局,基因修饰模式动物地位日益凸显。相较于野生型的模式动物,基因修饰模式动物能够对目标基因开展功能缺失或功能获得的研究,达到人类生理或病理更精确的模拟,因此更适合探索研究人类基因功能和人类疾病致病机制,是目前行业重要的发展方向。这其中Cre工具鼠和疾病模型鼠亦扮演着举足轻重的角色。一起围观下百奥动物的蓝宝小家伙带来的好物分享吧~ 鼠界“爱德华”--Cre工具鼠系列INTRODUCTION基因敲除鼠常被用来研究基因的功能,但据研究报道,小鼠有超过30%的基因敲除后导致了胚胎致死或者出生后不久死亡[2],从而无法对该基因在体内发挥的生理功能进行更好的研究。为了解决这个问题,科学家发现了Cre/loxP系统。 1 什么是Cre/loxP系统?Cre-loxp 系统来自于P1 噬菌体,是噬菌体感染病毒实现寄生的“杀手锏”。 Cre重组酶(cyclization recombination enzyme),它是一种由343个氨基酸组成的单体蛋白,可以引发loxP位点的DNA 重组。loxP是一段长34bp的DNA序列,是Cre重组酶识别的位点,被称为loxP位点。Cre重组酶的调控方式有多种多样,有如特异性基因启动子调控的Cre(Promoter-regulated Cre)、配体或药物诱导的Cre(Inducible Cre)、荧光报告的Cre(Fluorescent Cre)以及同时被不同启动子调控的Cre(Split Cre,NCre 和CCre)等。不同调控方式的选择造就了不同的Cre工具鼠。 2 Cre/loxP系统在基因编辑大小鼠中的应用根据Cre-loxp 系统工作的原理,在基因编辑大小鼠中,分别制作Cre重组酶工具大小鼠和带有loxp位点的flox鼠,通过杂交可以实现组织特异性基因敲除,这样可以避免全身性敲除引起的纯合致死情况。你是不是也正在经历实验设计选不到合适Cre鼠的窘境?百奥动物自主开发了一系列Cre工具鼠,助力不同应用场景的科学研究。B-Pgr-iCre小鼠基本信息应用领域生殖系统功能研究表型分析ROSA26-CAG-LSL-tdTomato mice是公司自主研发并验证成功的tdTomato报告小鼠品系。在与Cre小鼠交配后,剔除stop元件,可表达tdTomato红色荧光蛋白。本实验方案利用B-Pgr-iCre mice与CAG-LSL-tdTomato报告小鼠交配,iCre重组酶可介导子代小鼠中表达Pgr的细胞删除LSL元件,表达tdTomato红色荧光蛋白,荧光显微镜下可观察到红色荧光信号,可证明表达Pgr的细胞成功表达iCre重组酶并介导基因重组功能。上图为小鼠不同器官中实验结果的显示,(a)代表小鼠的生殖系统,包括子宫、输卵管和卵巢。(b)代表小鼠的乳腺。IF 检测 tdTomato 红色荧光蛋白和绿色荧光标记 Pgr(progesterone) 的表达。纵轴代表不同基因型的小鼠,横轴代表不同的荧光标记。B-Pgr-iCre(Mut/+);CAG-tdTomato(Mut/+) 小鼠表现出明显的 tdTomato 红色荧光蛋白表达。B-Cdh5-iCreERT2 mice小鼠基本信息应用领域心血管系统,肿瘤和细胞分化机制研究表型分析ROSA26-CAG-LSL-tdTomato mice是公司自主研发并验证成功的tdTomato报告小鼠品系。在与Cre小鼠交配后,剔除stop元件,可表达tdTomato红色荧光蛋白。本次实验中B-Cdh5-iCreERT2 mice与ROSA26-CAG-LSL-tdTomato mice交配,可实现Cdh5-iCreERT2(Mut/+);CAG-LSL-tdTomato(Mut/+)双阳性小鼠在tamoxifen诱导下,在血管淋巴管内皮细胞特异性表达tdTomato红色荧光蛋白。上图为双阳性小鼠主要器官中 tdTomato 和 CD31 的免疫荧光图像。CD31是内皮标记物。(比例尺:25 μm)百奥动物Cre工具鼠产品列表向上滑动查看列表 高能“cosplay”--疾病模型鼠系列INTRODUCTION疾病动物模型是研究人类疾病机制、诊断预后标志物发现、药物筛选和评价等的重要支撑条件。利用基因工程技术对基因进行修饰,可建立敏感动物品系和与人类疾病相同的疾病模型进行药物筛选和药效研究,这类动物可培育为稳定遗传的品系,且具有特定的病理特征,已经成为药物快速筛选的重要手段。百奥动物建立了丰富的疾病模型资源,为疾病治疗药物评价提供稳定有效的模型资源,并能够基于此为广大合作伙伴提供符合国际水平的药理药效服务。血友病模型--B-F8 KO 小鼠F8又称AHF,FVIII(coagulation factor VIII),是位于X染色体上编码凝血因子VIII的基因,在外源性和内源性凝血途径中都不可或缺。它作为凝血因子IXa的辅助因子参与凝血过程,同时借助于Ca2 +和膜磷脂,形成复合物以激活X因子,并进行一系列后续的凝血反应。F8基因的缺失导致严重的凝血障碍,形成伴X染色体隐性遗传的A型血友病。F8基因敲除模型为血友病患者的药物筛选提供了重要的临床前借鉴意义。模型数据验证将实验动物随机分组,分别尾静脉注射给予生理盐水或1mg/kg诺其,给药30min后腹主动脉采血检测血凝指标:活化部分凝血活酶时间 APTT。结果显示:F8 KO小鼠APTT值远高于WT小鼠,给予重组人凝血因子VIIa后,APTT恢复至正常值。结果证明:B-F8 KO小鼠可以作为抗凝血药药效验证的有力工具。高血糖和肥胖症模型--B-ob/ob 小鼠瘦素(Leptin)是由肽链构成的肽类激素。主要由脂肪细胞分泌,其表达主要在白色脂肪组织。此外,在心肌、骨骼肌、胎盘、肺、乳腺上皮和胃黏膜等均有表达。在功能上能够有抑制食欲,增加能量消耗,抑制脂肪合成促进其分解。胰岛素可促进瘦素的分泌,反过来瘦素对胰岛素的合成、分泌发挥负反馈调节。ob是瘦素的编码基因,百奥动物利用基因编辑技术将ob基因2、3号外显子敲除制备的B-ob/ob小鼠,具有肥胖症和高血糖症状,是研究高血糖和肥胖症的有力模型。模型数据验证纯合B-ob/ob小鼠4周龄后体重和血糖(-/-)均持续高于对照组。抗人GCGR抗体药物crotedumab在B-ob/ob小鼠体内的药效。(A)B-ob/ob小鼠的体重变化。(B-C)非空腹血糖和空腹血糖测量。将8-10周龄的雄性B-ob/ob小鼠随机分为2组,每组6-7只。给药组第0天进行给药,第0、3、7天测定非空腹血糖,禁食6小时后测定空腹血糖。(D)Crotedumab可改善葡萄糖耐量。(E)血糖含量曲线下面积。小鼠在自由取水的条件下禁食6h,尾尖取血测定空腹血糖(0 min),腹腔注射葡萄糖2 g/kg,在指定时间测定血糖。(F-G)胰高血糖素和胰岛素测量。结果显示,crotedumab在禁食和非禁食状态下均具有降糖作用,同时能够改善B-ob/ob小鼠的葡萄糖耐量。百奥动物疾病模型鼠产品列表百奥动物可提供多种稳定优质的基因编辑自发疾病动物模型,如肿瘤模型鼠(B-p53 KO 小鼠,B-p53 KO大鼠),高血糖模型鼠(B-ob/ob小鼠),凝血模型鼠(B-F8 KO小鼠,B-F9 KO小鼠)等,提供客户用于相关研究。向上滑动查看产品列表意犹未尽?想要共享蓝宝的更多好物清单,欢迎扫描下方二维码查看或者来电咨询。参考资料:[1]https://mp.weixin.qq.com/s/rhhEFL5A9KhVKAULO29COw[2]Huimin Zhang, Qi Zheng, Ruby Yanru Chen-Tsai;Establishment of a Cre-rat resource for creating conditional and physiological relevant models of human diseases. Transgenic Res. 2021 Feb;30(1):91-104
近年来,免疫治疗在抗肿瘤治疗中的作用备受关注,基于免疫治疗的新药开发和标志物探索,成为了当前肿瘤研究的热点,这也对临床前研究动物模型的建立提出了更高的要求。即可以模拟人肿瘤特征又同时存在“人源化”免疫系统的免疫重建小鼠模型,成为了免疫肿瘤研发中的优质模型。百奥动物B-NDG小鼠缺乏成熟的T、B、NK细胞,是目前国际公认的免疫缺陷程度高、适合人源细胞或组织移植的工具小鼠。将人的免疫细胞、造血干细胞移植到B-NDG及B-NDG衍生小鼠中构建的免疫系统重建小鼠,能够更好的模拟人的免疫系统,进行免疫学研究和免疫药物评价。但在使用重度免疫缺陷小鼠进行PBMC和CD34+ HSC免疫重建时会面临一些常见问题,如PBMC重建后的T细胞会对小鼠自身细胞进行攻击导致严重的xeno-GvHD反应;CD34+ HSC重建后的NK细胞和髓系细胞重建比例不足。我们以B-NDG小鼠为基础分别开发了可延缓xeno-GvHD反应和可促进髓系细胞发育的二代系列小鼠,以满足特定细胞功能研究和相应靶点药物评价的需求。减轻GvHD反应,延长实验窗口期B-NDG MHC I/II DKO mice plus 不表达MHC I/II 类分子B-NDG小鼠、B-NDG B2m KO plus小鼠和B-NDG MHC I/II DKO plus小鼠进行人PBMC重建后GvHD严重程度的比较 第0天将3个健康供体的人PBMCs (5E6)静脉注射B-NDG小鼠、B-NDG B2m KO plus小鼠和B-NDG MHC I/II DKO plus小鼠(雌性,5周龄,n=5)中。A.用Kaplan Meier生存曲线分析小鼠存活率。B.体重变化。GvHD临床体征每周评分两次。结果表明:B-NDG MHC I/II DKO plus小鼠可显著延缓GvHD发生,并降低GvHD严重程度。因此B-NDG MHC I/II DKO plus小鼠更适合将人PBMC移植到免疫缺陷小鼠模型。B-NDG B2m KO mice plus B-NDG小鼠、B-NDG B2m KO plus进行人PBMC重建后GvHD严重程度的比较将3个健康供体的2E6人PBMC静脉注射B-NDG B2m KO plus小鼠(雌性,11周龄,n=5)和B-NDG小鼠(雌性,10周龄,n=6)中。GvHD临床体征每周评分两次。结果表明:在人PBMC诱导的GvHD模型中,B-NDG B2m KO plus小鼠比B-NDG小鼠生存期明显延长,B-NDG B2m KO plus小鼠可延缓GvHD的发病并减轻其严重程度。促进髓系细胞重建B-NDG MGMT3 mice 人源化IL3, CSF2, CSF1, THPO 基因未经辐照的B-NDG MGMT3小鼠进行人CD34+HSC重建将人CD34+HSC造血干细胞(3E4)静脉注射B-NDG小鼠和B-NDG MGMT3小鼠(出生后24-72h,n=15)。B-NDG小鼠给予1.0 gy辐照,B-NDG MGMT3小鼠不辐照。流式细胞分析两种小鼠进行人CD34+HSC免疫重建后的外周血淋巴细胞。结果表明:未经辐照的B-NDG MGMT3小鼠的CD45+细胞比例从移植后12周开始达到25%,并持续上升,明显高于B-NDG小鼠。B-NDG MGMT3小鼠单核细胞、MDSCs、DCs和Treg的比例高于B-NDG小鼠。(注:A面板第18周数据因流式细胞检测问题无意义。)B-NDG hCSF1/hTHPO mice 未经辐照的B-NDG hCSF1/hTHPO小鼠进行人CD34+HSC重建将人CD34+HSC细胞(0.15 M)静脉注射到纯合B-NDG hCSF1/hTHPO小鼠(雌性,6周龄,n=15)。流式细胞术分析人CD34+HSC免疫重建后小鼠外周血淋巴细胞。结果显示:未经辐照的B-NDG hCSF1/hTHPO小鼠成功重建了T、包括B、NK、髓系细胞、单核细胞和中性粒细胞在内的人多系细胞。免疫缺陷动物产品列表
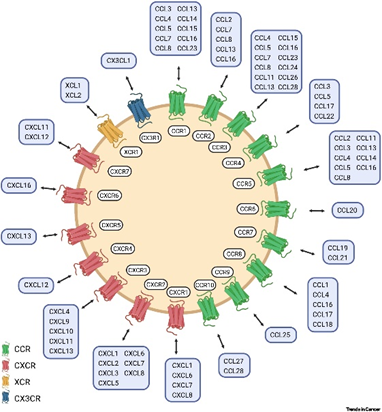
导读趋化因子是一类分子量小的细胞因子,其主要作用是在稳态和病理条件下募集白细胞亚群,又被称为趋化性细胞因子。根据其主要蛋白质结构的前两个半胱氨酸(C)残基的位置,将趋化因子分为 C 、CC 、CXC 和 CX3C 趋化因子四大亚家族,其主要负责参与调控机体的器官发育、免疫监视、宿主防御和组织更新等生理过程。趋化因子也可以根据其表达和功能分为炎性趋化因子和稳态趋化因子。炎症性细胞因子在炎症部位迅速分泌,从而将效应细胞募集到发炎组织中;而稳态趋化因子在生理条件下组成性表达并在细胞迁移和归巢中发挥作用,因此趋化因子在协调炎症及正常状态下的体内细胞群定位中发挥核心作用。 图1.趋化因子配体与受体[1]趋化因子受体表达于细胞表面,是与G蛋白偶联的7次跨膜蛋白,趋化因子就是与受体结合后传递细胞信号的,故受体根据其结合的趋化因子亚家族来命名,如 XCR、CCR、CXCR、CX3CR 等。这些受体负责调控多条细胞信号通路,如调动肌动蛋白聚合、细胞骨架重排、粘着斑组装和解聚,此外也在细胞存活等生命活动中发挥着重要作用。图2.趋化因子信号通路图[2]此外,趋化因子还参与多种癌症发展过程,如血管生成、转移、癌细胞增殖、干性和侵袭性,是疾病进展的关键决定因素,对治疗反应和患者预后有很大影响。由于它们在癌细胞和免疫浸润细胞中重要的调节功能,使得趋化因子配体及其受体成为非常强大的治疗靶点。趋化因子的靶向治疗目前,国内外已有多家药企开启了针对趋化因子及其受体的药物开发,其中临床已批准的靶向趋化因子的药物包括:2012年上市的抗CCR4抗体(Mogamulizumab)和2007年上市的CXCR4拮抗剂(Maraviroc)等,用于治疗恶性血液瘤。此外,还有更多的针对不同趋化因子受体-配体轴作为癌症治疗策略的多种努力,这些治疗策略目前已表现出巨大的潜力,正处于临床开发中。数据来源于科睿唯安趋化因子及其受体对于维持机体稳态具有重要意义。然而一旦趋化因子活性失控,则会导致慢性炎症和自身免疫性疾病。针对一系列疾病靶点的研究机理,BioMice百奥动物自主开发了CCR家族靶点人源化小鼠,包括CCR1-CCR9,可以为该靶点药物的开发提供有效的临床前药效评价工具,助力靶向药物研究。B-hCCR1 mice基本信息 蛋白表达分析 通过流式细胞术检测野生型C57BL/6和纯合B-hCCR1小鼠中CCR1的蛋白表达。收集野生型C57BL/6和纯合B-hCCR1小鼠的腹腔巨噬细胞,并用种属特异性抗CCR1抗体进行流式细胞术分析。结果显示:鼠CCR1仅在野生小鼠中检测到,人CCR1仅在纯合B-hCCR1小鼠中检测到。 B-hCCR2 mice基本信息 蛋白表达分析 通过流式细胞术检测野生型C57BL/6和纯合B-hCCR2小鼠中CCR2的蛋白表达。收集野生型C57BL/6和纯合B-hCCR2小鼠的骨髓,并用种属特异性抗CCR2抗体进行流式细胞术分析。结果显示:鼠CCR2仅在野生小鼠中检测到,人CCR2仅在纯合B-hCCR2小鼠中检测到。 B-hCCR3 mice基本信息 蛋白表达分析 通过流式细胞术检测野生型C57BL/6和纯合B-hCCR3小鼠中CCR3的蛋白表达。收集野生型C57BL/6和纯合B-hCCR3小鼠的骨髓,并用种属特异性抗CCR3抗体进行流式细胞术分析。结果显示:鼠CCR3仅在野生小鼠中检测到,人CCR3仅在纯合B-hCCR3小鼠中检测到。 B-hCCR4 mice基本信息 蛋白表达分析 通过流式细胞术检测野生型C57BL/6和纯合B-hCCR4小鼠中CCR4的蛋白表达。收集野生型C57BL/6和纯合B-hCCR4小鼠的脾细胞,并用种属特异性抗CCR4抗体进行流式细胞术分析。结果显示:鼠CCR4仅在野生小鼠中检测到,人CCR4仅在纯合B-hCCR4小鼠中检测到。 B-hCCR4小鼠的T细胞能与抗人CCR4抗体结合通过流式细胞术(FACS)分析B-hCCR4小鼠的T细胞结合抗人CCR4抗体的能力。收集B-hCCR4 小鼠的脾细胞(雌性,6周龄),使用FACS检测T细胞与抗人CCR4抗体(内部合成)的结合。结果显示:与同型对照相比,B-hCCR4小鼠的T细胞可以很好地结合抗人CCR4抗体。 B-hCCR5 mice基本信息 蛋白表达分析 通过流式细胞术检测野生型C57BL/6和纯合B-hCCR5小鼠中CCR5的蛋白表达。收集野生型C57BL/6和纯合B-hCCR5小鼠的腹腔冲洗液,并用种属特异性抗CCR5抗体进行流式细胞术分析。结果显示:鼠CCR5仅在野生小鼠中检测到,人CCR5仅在纯合B-hCCR5小鼠中检测到。 B-hCCR6 mice基本信息 蛋白表达分析 通过流式细胞术检测野生型C57BL/6和纯合B-hCCR6小鼠中CCR6的蛋白表达。收集野生型C57BL/6和纯合B-hCCR6小鼠的脾细胞,并用种属特异性抗CCR6抗体进行流式细胞术分析。结果显示:鼠CCR6仅在野生小鼠中检测到,人CCR6仅在纯合B-hCCR6小鼠中检测到。B-hCCR7 mice基本信息蛋白表达分析 通过流式细胞术检测野生型C57BL/6和纯合B-hCCR7小鼠中CCR7的蛋白表达。收集野生型C57BL/6和纯合B-hCCR7小鼠的脾细胞,并用种属特异性抗CCR7抗体进行流式细胞术分析。结果显示:鼠CCR7仅在野生小鼠中检测到,人CCR7仅在纯合B-hCCR7小鼠中检测到。 B-hCCR8 mice基本信息 蛋白表达分析 人CCR8在纯合B-hCCR8小鼠肿瘤中CD4+ T细胞和Treg细胞均可检测到,但在脾细胞和血细胞中不能检测到。鼠CCR8在野生型小鼠的肿瘤中可检测到,脾细胞中弱表达,而在血细胞中不表达。血常规检测 收集野生型C57BL/6和B-hCCR8小鼠的外周血进行血常规检测(n=8,雌性,9周龄),结果显示:B-hCCR8小鼠的各指标与野生C57BL/6小鼠无明显差异,表明CCR8的人源化未改变血细胞的组成及形态。血生化检测收集野生型C57BL/6和B-hCCR8小鼠的血清进行血生化检测(n=8,雌性,9周龄),结果显示:B-hCCR8小鼠的各指标与野生C57BL/6小鼠无明显差异,表明CCR8的人源化未影响小鼠的肝、肾功能及脂肪代谢能力。 抗人CCR8抗体药效验证 抗人CCR8抗体在B-hCCR8小鼠接种MC38模型和B-Tg(hCCL1) MC38模型中均有较好的抑瘤效果。TILs分析显示:抗人CCR8抗体给药组(G2、G4)与未给药组(G1、G3)相比,总的Tregs和hCCR8+ Tregs的比例显著性降低。 B-hCCR9 mice基本信息 蛋白表达分析 通过流式细胞术检测野生型C57BL/6和纯合B-hCCR9小鼠中CCR9的蛋白表达。收集野生型C57BL/6和纯合B-hCCR9小鼠的胸腺细胞,并用种属特异性抗CCR9抗体进行流式细胞术分析。结果显示:鼠CCR9仅在野生小鼠中检测到,人CCR9仅在纯合B-hCCR9小鼠中检测到。趋化因子受体及其配体的相互作用非常复杂,尽管面临挑战,但目前还是有大量针对不同趋化因子受体的抑制剂正在临床前研究或临床试验阶段,可以相信未来趋化因子受体抑制剂将在调节TME的组成并优化患者的免疫反应方面发挥重大作用,为肿瘤患者带来更多希望。参考文献[1] Märkl, F., Huynh, D., Endres, S. & Kobold, S. Utilizing chemokines in cancer immunotherapy. Trends in Cancer 8, 670–682 (2022)[2] R&D Systems.lnc.The Chemokine Superfamily: Critical Regulators of Homeostasis & Inflammation,2019
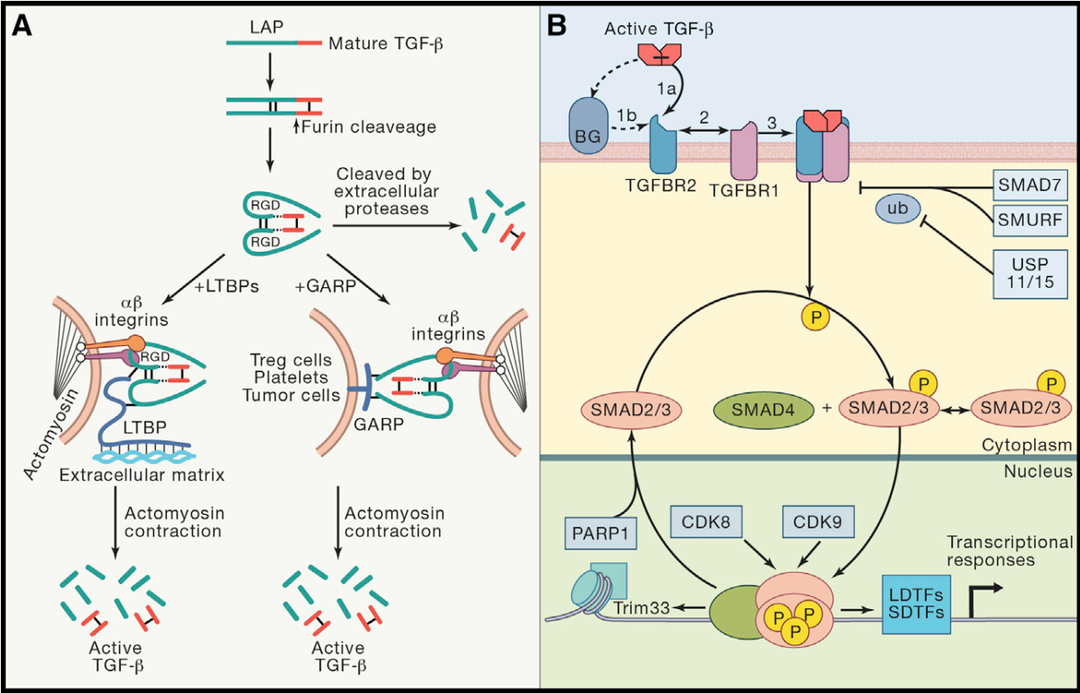
转化生长因子β(TGFβ)是一种多功能的细胞因子,对细胞增殖、分化、粘附、迁移和凋亡具有多重功能。多种类型细胞均可分泌,最早从人血小板中鉴定。哺乳动物中表达三种类型TGFβ:TGFβ1、TGFβ2和TGFβ3,每种由不同的基因编码,并通过结合相同的TGFβ受体发挥相应功能。其中TGFβ2和TGFβ3在胚胎发育中发挥重要作用,而TGFβ1则主要参与控制出生后的免疫反应[1]。三种TGFβ起初均表达为非活性蛋白复合物(SLC或LLC)。TGFβ1同型二聚体与潜伏相关肽(Latent Associated Peptide, LAP)(TGFβ1 N 端具有的长 20~30 个氨基酸的序列)形成的复合被称为小潜复合物 (Small Latent Complex, SLC)。在细胞外,SLC 与 潜伏TGFβ1结合蛋白(Latent TGFβ Binding Protein, LTBP)结合形成的复杂复合物被称为大潜复合物(Large Latent Complex, LLC)。TGFβ的活化有以下3种途径:1)SLC在胞外被蛋白酶水解;2)LLC被细胞外基质铆钉,进而由αβ整合素介导释放有活性的TGFβ;3)SLC被细胞表面的GARP铆钉,并由αβ整合素介导释放有活性的TGFβ。接下来,具有活性的TGFβ与TGFβRII二聚体结合,随后进一步与TGFβRI二聚体结合,形成四聚体,激活下游信号通路,调节基因表达。图1. TGFβ信号通路[2,3]稳态条件下,TGFβ1信号不仅调节多种组织细胞的生长、增殖和分化;也指导免疫系统的耐受和炎症抑制,尤其在胃肠道起重要作用。TGFβ1调节功能的多样性赋予了它在肿瘤发生和发展中亦正亦邪的双面性。在癌症发生早期,TGFβ是一个抑癌因子,可发挥细胞周期阻滞作用,阻断细胞从G1期进入S期,达到抑制肿瘤细胞增殖的作用。然而,随着炎症加剧,一些癌细胞可能会发生突变,使其对TGFβ信号通路不响应,或响应与凋亡脱钩、反变为促进癌细胞转移和定植。同时,肿瘤细胞亦可大量分泌TGFβ1发挥免疫抑制功能,抑制T、B淋巴细胞与NK细胞活化和分化,造成机体免疫功能障碍,使得肿瘤细胞免疫逃逸(图2)。图2. TGFβ的抑癌和促癌信号[2]针对TGFβ信号通路的在研药物主要包括配体抗体、配体陷阱(如bintrafusp alfa)、受配体抗体、小分子激酶抑制剂等,主要聚焦于肿瘤免疫疗法[4]。目前,处于临床阶段的TGFβ1配体抗体药物如下表所示:数据整理自Cortellis数据库为了更好的助力靶向TGFβ1的新药研发,百奥动物自主研发了TGFβ1人源化小鼠B-hTGFB1 mice,用于靶向TGFβ1药物的临床前药效评估。B-hTGFB1 mice 图3. RT-qPCR分析B-hTGFB1小鼠和野生型小鼠的TGFB1 mRNA的表达。B-hTGFB1小鼠中人TGFB1 mRNA表达量与野生型小鼠中的鼠TGFB1 mRNA表达量类似。这表明人TGFB1基因原位替换不影响TGFB1 mRNA的表达。图4. 利用流式细胞仪分析纯合B-hTGFB1小鼠和野生型小鼠的血小板中TGFB1蛋白的种属特异性表达。小鼠的TGFB1仅在野生型小鼠中被检测到,人TGFB1仅在纯合B-hTGFB1小鼠中检测到。脾脏中TGFB1蛋白的表达同样也具有种属特异性(数据未展示)。TGFB1人源化不影响脾脏、淋巴结、血液中白细胞各亚群比例(数据未展示)。图5. 抗体结合实验。从B-hTGFB1小鼠和野生型小鼠取血,用流式细胞术分析血小板人LAP的表达。SRK-181可特异性结合SLC并抑制TGFβ1活化[5]。于是,在SRK-181处理过的纯合B-hTGFB1小鼠和野生型小鼠中均能检测到人LAP的表达;而未处理的鼠中不能检测到人LAP的表达。(hLAP抗体可人鼠交叉识别。)图6. 抗鼠PD-1抗体和抗人TGFβ1抗体联用在B-hTGFB1小鼠中的抗肿瘤作用。(A)mPD-1和hTGFβ1抗体(自制)联用可抑制B-hTGFB1小鼠中MC38肿瘤的生长;(B)治疗期间的体重改变。数值为平均值±SEM。由图可见,抗鼠PD-1抗体和抗人TGFβ1抗体联用可有效控制B-hTGFB1小鼠中的肿瘤生长,说明B-hTGFB1小鼠是TGFβ1抗体药物临床前评估的有力模型。相关产品列表 品系货号B-hTGFB1 mice112245B-hTGFBR2 mice110874B-hGARP mice110102B-hGARP/hTGFB1 mice112241B-hLRRC33 mice110757B-Tgfβ1 cKO mice110164参考资料1. Stockis, J., Dedobbeleer, O. & Lucas, S. Role of GARP in the activation of latent TGF-β1. Molecular bioSystems 13, 1925-1935 (2017).2. Batlle, Eduard, and Joan Massagué. “Transforming Growth Factor-β Signaling in Immunity and Cancer.” Immunity vol. 50,4 (2019): 924-940. doi:10.1016/j.immuni.2019.03.0243. Kelly, Aoife et al. “Regulation of Innate and Adaptive Immunity by TGFβ.” Advances in immunology vol. 134 (2017): 137-233. doi:10.1016/bs.ai.2017.01.0014. Kim, Byung-Gyu et al. “Novel therapies emerging in oncology to target the TGF-β pathway.” Journal of hematology & oncology vol. 14,1 55. 6 Apr. 2021, doi:10.1186/s13045-021-01053-x5. Martin, Constance J et al. “Selective inhibition of TGFβ1 activation overcomes primary resistance to checkpoint blockade therapy by altering tumor immune landscape.” Science translational medicine vol. 12,536 (2020): eaay8456. doi:10.1126/scitranslmed.aay8456
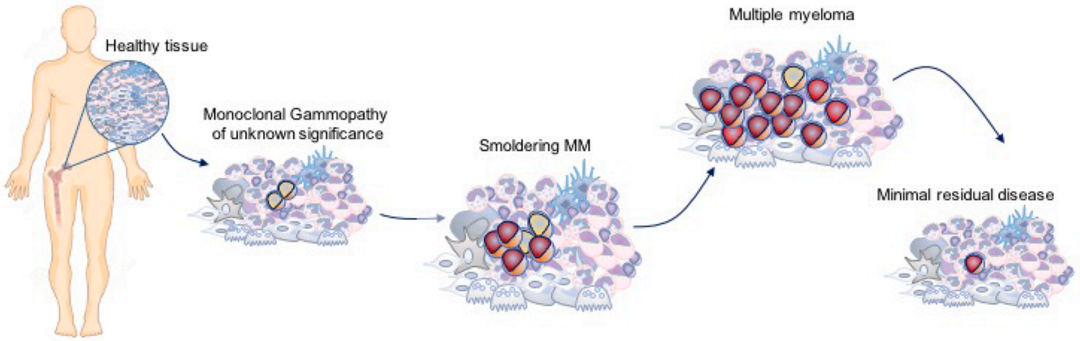
多发性骨髓瘤(Multiple Myeloma, MM)是全球第二大常见的恶性血液肿瘤,始于骨髓中健康的浆细胞恶性增殖。在当前是一种较难治愈的疾病,大多数患者终将复发。据统计,2020年全球有超过17万人被诊断出患有多发性骨髓瘤,60岁以上老人属于该病的高发群体,发病年龄亦有呈年轻化的趋势。随着我国人口老龄化趋势上升,多发性骨髓瘤的防治负担将逐年加剧,成为危害人们身体健康的一大挑战。多发性骨髓瘤从癌前到有明显疾病症状的演变过程,在SMM阶段的早期发现和早期干预以及预防或治疗策略可能是提高这种复杂疾病治愈率的途径。[1] 左图:正常的骨髓 ,右图:多发性骨髓瘤;多发性骨髓瘤中可见大量恶性浆细胞,其特征是在细胞核附近的细胞质内有一个苍白的区域。[2]目前,针对MM的治疗药物有糖皮质激素、细胞毒性药物、免疫抑制剂、蛋白酶抑制剂、单抗和细胞疗法等。其中,免疫治疗已经在许多癌症领域被证明是革命性疗法,但由于MM存在免疫抑制微环境现象,影响了免疫治疗的疗效,导致其对MM的治疗进展较为缓慢。MM细胞与骨髓(BM)微环境之间的相互作用,对MM的发病起着关键作用,可通过诱导或分泌细胞因子促进肿瘤细胞生长、免疫逃逸及耐药,增加Treg细胞的数量,抑制效应T细胞的杀伤等。靶向特定肿瘤抗原或逆转具有免疫抑制作用的骨髓微环境的免疫疗法可以帮助改善MM的标准治疗方案。骨髓MM微环境中的抗骨髓瘤作用[3]细胞重定向BiAb和BiTE同时结合MM细胞上的骨髓瘤特异性抗原和T细胞上的CD3。MM抗原包括BCMA, CD38, CS1/SLAMF7, GPRC5D和FcRH5,如图示。BiAbs或自然杀伤细胞衔接器(NKCEs)也靶向自然杀伤(NK)细胞相关受体抗原(如CD16A, NKG2D, NKp30),激活NK细胞并增强其抗MM活性。双特异性分子,包括双特异性抗体(BsAbs)和双特异性T细胞衔接器(BiTEs),通过同时结合MM细胞和免疫效应细胞上的抗原,使这些细胞靠近从而促进免疫细胞裂解MM细胞。靶向BCMA和GPRC5D的BsAbs已展示出很好的临床疗效,针对FcRH5的早期临床试验结果也非常有前景。这些药物的免疫调节作用不依赖主要组织相容性复合体(MHC) I类的抗原提呈,可在没有共刺激的情况下发生,适用于免疫系统功能失调的MM患者。BCMA01全称B细胞成熟抗原,又名CD269或TNFRSF17,属于肿瘤坏死因子受体超家族成员,仅高表达于浆细胞表面,部分表达于浆细胞样树突状细胞,是MM免疫治疗的理想靶点。针对BCMA靶点开发的肿瘤免疫疗法主要分为3类:嵌合抗原受体T细胞疗法(CAR-T)、双特异性抗体(BsAb)、抗体药物偶联物(ADC)。GPRC5D02全称G蛋白偶联受体C57家族亚型D,为7次跨膜蛋白,高表达于浆细胞表面,低表达于毛囊区域,其他健康细胞则不表达,为治疗MM的潜在候选靶点。GPRC5D的表达与BCMA不相关,联合靶向这两个靶点的疗法可以发挥互补效应或开发双靶点CAR-T、双抗。FcRH503又称FcRL5、CD307或免疫球蛋白超家族受体易位相关蛋白2 (Immunoglobulin Superfamily Receptor Translocation Associated 2, IRTA2),是一种功能未知的膜蛋白,只表达于B细胞系,包括骨髓瘤细胞。临床前研究的数据表明,FcRH5/CD3双抗可成功激活T细胞,诱导细胞因子产生,并清除恶性浆细胞。根据科睿唯安数据库检索,靶向MM相关靶点药物研究数量众多,部分靶点研究进展情况见下表。强生的Darzalex是在2015年第一批被批准用于MM治疗的免疫疗法,这一时期MM的抗体药研发主要集中在靶向CD38靶点;到2020年GSK研发的首个靶向BCMA药物Blenrep在美获批,2021年BMS的Abecma上市,今年传奇生物的Carvykti上市以及强生的Teclistamab作为全球首个CD3/BCMA双抗获批即将上市,另外还有众多管线处在临床前或临床试验阶段,靶向BCMA赛道的药物研发可谓相当火热。布局靶向MM治疗药物的新靶点,避免同质化竞争,或许也不失为后来者开发该适应症药物以占得先机的上策,当然面临的风险也随之提高。部分药物研究进展整理自科睿唯安数据库及网络百奥动物利用基因编辑技术自主开发了MM相关靶点人源化小鼠及细胞系,助力抗体药物临床前研究。部分数据展示如下:B-hCD38 mice蛋白表达分析采集野生型C57BL/6小鼠和纯合B-hCD38小鼠的脾细胞和血液,用种特异性抗CD38抗体进行流式细胞术分析。小鼠CD38在WT小鼠中检测到。纯合B-hCD38中只检测到人CD38,而WT小鼠检测不到人CD38。药效验证将小鼠T淋巴细胞瘤B-hCD38-luc E.G7-OVA细胞经尾静脉注射到B-hCD38纯合小鼠(雌性,6周龄,n=6)体内。当总通量达到约106 Ig时,将小鼠分组,并用抗人CD38抗体对其进行治疗。(A) 抗人CD38抗体(内部合成)抑制B-hCD38-luc E.G7-OVA小鼠肿瘤生长。(B) 治疗期间体重变化。(C) B-hCD38-luc E.G7-OVA细胞的体内荧光素酶成像图。每周2次测量信号强度和体重,第0、3、7、10天行影像学检查。值表示为平均值±SEM。B-hBCMA micemRNA表达分析RT-PCR分析野生型C57BL/6小鼠和B-hBCMA小鼠BCMA基因特异性表达情况,鼠Bcma mRNA仅在野生型C57BL/6小鼠脾细胞中检测到,人BCMA mRNA仅在纯合B-hBCMA小鼠中检测到。B-hGPRC5D micemRNA和蛋白表达分析(A) RT-PCR分析野生型C57BL/6小鼠和B-hGPRC5D小鼠GPRC5D基因的特异性表达情况,鼠Gprc5d mRNA仅在野生型C57BL/6小鼠睾丸中检测到。人GPRC5D mRNA仅在纯合B-hGPRC5D小鼠中检测到。(B) 取野生型C57BL/6小鼠和纯合B-hGPRC5D小鼠的脾脏,用抗GPRC5D抗体进行western blot分析。由于抗体的交叉反应性,GPRC5D在WT小鼠和纯合B-hGPRC5D小鼠中均可检测到。 相关产品列表 更多数据信息,欢迎联系我们。参考资料[1] https://doi.org/10.1016/j.ctrv.2021.102284[2] https://www.cancer.net/cancer-types/multiple-myeloma/introduction[3] doi: 10.3389/fonc.2022.1032775[4] doi:10.3390/jcm9072166

趋化因子(chemokines)是一类由细胞分泌的小细胞因子或信号蛋白,具有诱导附近反应细胞定向趋化的能力,通过与G蛋白偶联跨膜受体(称为趋化因子受体,选择性地表达在靶细胞表面)相互作用来发挥其生物学效应。根据结构(氨基端(N端)半胱氨酸的排列方式),趋化因子被分为四个主要亚家族:CXC(CXCL1-17)、CC(CCL1-28)、XC(XCL1、XCL2)和CX3C(CX3CL1)四个亚族。 CC趋化因子亚族又称β趋化因子亚家族,其氨基末端有2个相近的半胱氨基结构。CC趋化因子亚族包含27种类型,依次为CCL1-CCL28(其中CCL10与CCL9相同),是一类可引起免疫细胞定向迁移至炎症部位的小分子分泌性蛋白。常见的有嗜酸性粒细胞趋化因子、单核细胞趋化蛋白及其他细胞亚群的趋化因子等。其受体有10个,分别为CCR1-CCR10。CC型趋化因子可趋化淋巴细胞、单核细胞、酸性粒细胞等细胞的游走趋化,在炎症反应、清除抗原、细菌感染等方面起着重要作用。CC趋化因子亚族列表CC趋化因子具有促癌和抗癌双重特性,主要对中性粒细胞、单核细胞、肥大细胞、树突细胞、NK细胞、T和B淋巴细胞等具有强大的趋化活性。比较重要的有:单核细胞趋化蛋白(MCP-1/CCL2)、巨噬细胞炎症蛋白(MIP/CCL3)、RANTES/CCL5等。01CCL2在肿瘤侵袭转移中的作用 CCL2在肿瘤微环境中由肿瘤细胞和基质细胞表达,在肿瘤原发部位诱导肿瘤细胞增殖,刺激肿瘤细胞向周围细胞外基质迁移和侵袭,随后CCL2促进肿瘤细胞内渗进入循环,可能是通过招募宿主骨髓细胞来促进这一过程。一旦进入血液循环,CCL2可能会引导癌细胞沿着趋化梯度向转移部位扩散。肿瘤细胞被困在小毛细血管中会引发肿瘤细胞外渗,这种外渗进一步受到CCR2+髓系细胞和CCR2+内皮细胞的支持。最后,CCL2通过募集更多的骨髓细胞和内皮细胞促进转移部位的肿瘤生长和肿瘤定植。[1]02CCL3在白血病骨髓微环境中的作用 在白血病BM微环境(蓝色阴影区域)中,CCL3可诱导支持白血病细胞优势增殖的多个过程:(1)正常龛细胞向白血病适应细胞的转化;(2)选择性抑制正常HSPCs;(3)从BM中动员正常HSPCs。缩写:BM,骨髓;HSPC,造血干/祖细胞。[2]03CCL20在癌症中的作用 CCL20-CCR6信号通路通过增强癌细胞的迁移和增殖直接促进癌症进展,通过免疫细胞控制重塑肿瘤微环境间接促进癌症进展。[3]靶向趋化因子及其受体治疗人类肿瘤、自身免疫性疾病和慢性炎症愈发受到重视。然而,到目前为止针对趋化因子和趋化因子受体的药物研发才刚刚起步。已获批的抗体药物有CCR4抗体(Mogamulizumab)和CXCL8(IL-8)抗体ABCream。为了推动靶向趋化因子的药物开发,百奥动物自主研发了一系列趋化因子人源化动物&细胞模型。B-hCCL2 mice蛋白表达分析ELISA法检测纯合B-hCCL2小鼠种属特异性CCL2表达分析。收集野生型小鼠和纯合B-hCCL2小鼠体内抗mCD3ε抗体刺激4h的血清,用种属特异性CCL2 ELISA试剂盒进行ELISA分析。小鼠CCL2仅在野生型小鼠中可检测到。人CCL2仅在纯合B-hCCL2小鼠中可检测到,但在野生型小鼠中未检测到。B-hCCL3 mice蛋白表达分析ELISA法检测纯合B-hCCL3小鼠种属特异性CCL3表达分析。从野生型小鼠和纯合B-hCCL3小鼠中分离骨髓源性巨噬细胞(BMDM),体外用10 ng/mL LPS刺激,用种属特异性CCL3 ELISA试剂盒ELISA分析细胞培养上清液。在野生型小鼠中可检测到小鼠CCL3。人CCL3仅在纯合B-hCCL3小鼠中可检测到,但在野生型小鼠中未检测到。B-hCCL20 mice蛋白表达分析ELISA法检测纯合B-hCCL20小鼠中种属特异性CCL20表达分析。收集野生型小鼠和纯合B-hCCL20小鼠胸腺研磨上清,用种属特异性CCL20 ELISA试剂盒进行ELISA分析。在野生型小鼠中可检测到小鼠CCL20。人CCL20仅在纯合B-hCCL20小鼠中可检测到,但在野生型小鼠中未检测到。B-hCCL22 mice蛋白表达分析ELISA法对野生型小鼠和B-hCCL22小鼠中CCL22表达进行种属特异性分析。收集野生型小鼠和纯合B-hCCL22小鼠胸腺匀浆,用种属特异性CCL22 ELISA试剂盒进行ELISA分析。在野生型小鼠中可检测到小鼠CCL22。人CCL22 仅在纯合B-hCCL22小鼠中可检测到,但在野生型小鼠中未检测到。数值表示为平均值±SEM。ND:未检出。B-hCCR8/hCCL1 mice肿瘤微环境中CCL1的检测ELISA法检测C57BL/6和B-hCCR8/hCCL1荷瘤MC38细胞小鼠肿瘤微环境中CCL1。将小鼠结肠癌MC38细胞皮下植入C57BL/6和B-hCCR8/hCCL1小鼠(n=2或3)。当肿瘤体积约为600mm3时取肿瘤组织,并通过ELISA进行分析。在纯合B-hCCR8/hCCL1小鼠中可检测到人CCL1,含量约为150 pg/mg总蛋白。不同小鼠肿瘤浸润淋巴细胞的比较不同小鼠的肿瘤浸润淋巴细胞分析。B-hCCR8/hCCL1小鼠Treg细胞百分比(Th细胞%)与C57BL/6小鼠和B-hCCR8小鼠相比无显著差异。B-hCCL2 MC38蛋白表达分析ELISA法对B-hCCL2 MC38细胞中CCL2的表达进行分析。在B-hCCL2 MC38细胞的上清液中检测到人CCL2,但在野生型MC38细胞中未检测到。在野生型MC38细胞上清液中未检测到小鼠Ccl2。使用B-hCCL2 MC38细胞的2-D07克隆进行体内实验。肿瘤生长曲线和体重变化B-hCCL2 MC38细胞皮下同种移植瘤生长。将B-hCCL2 MC38细胞(5x105)和野生型MC38细胞(5x105)皮下植入C57BL/6N小鼠(雌性,7周龄,n=5)。每周测量两次肿瘤体积和体重。(A)平均肿瘤体积±SEM。(B)体重(平均值±SEM)。体积用mm3表示,公式为:V=0.5×长直径×短直径2。如A图所示,B-hCCL2 MC38细胞能够在体内建立肿瘤,并可用于疗效研究。B-Tg(hCCL1) MC38肿瘤浸润淋巴细胞分析与MC38细胞相比,荷瘤B-Tg(hCCL1)MC38细胞中CD3+ T细胞、CD4+ T细胞和Treg细胞占CD45+细胞的百分比显著增加。数据表示为平均值±SEM,采用单因素方差分析,然后进行Tukey检验,与其他列进行比较(*p<0.05,**p<0.01,***p<0.001,****p<0.0001)肿瘤生长曲线和体重变化B-Tg(hCCL1) MC38细胞皮下同种移植物肿瘤生长。B-Tg(hCCL1) MC38细胞(5x105)和野生型MC38细胞(5x105)皮下植入B-hCCR8小鼠(雌性,8周龄,n=7)。每周测量两次肿瘤体积和体重。(A)平均肿瘤体积±SEM。(B)体重(平均值±SEM)。体积用mm3表示,公式V=0.5×长径×短径2。如A图所示,B-Tg(hCCL1) MC38细胞能够在体内建立肿瘤,并可用于疗效研究。B-Tg(mCcl2) MC38肿瘤生长曲线和体重变化B-Tg(mCcl2) MC38细胞皮下同种移植肿瘤生长。将B-Tg(mCcl2) MC38细胞(5x105)和野生型MC38细胞(5x105)皮下植入C57BL/6小鼠(雌性,6周龄,n=5)。每周测量两次肿瘤体积和体重。(A)平均肿瘤体积±SEM。(B)体重(平均值±SEM)。体积用mm3表示,公式为:V=0.5×长直径×短直径2。如A图所示,B-Tg(mCcl2) MC38细胞能够在体内形成肿瘤,可用于疗效研究。肿瘤中mCcl2的表达分析实验结束时采集肿瘤细胞,ELISA检测小鼠Ccl2表达。如图所示,小鼠Ccl2在肿瘤匀浆中高表达。数据表示为平均值±SEM,采用T检验进行分析并与G1进行比较。(*p<0.05,**p<0.01,***p<0.001,****p<0.0001)参考资料1、Lim, Su et al. Oncotarget. 7 (2016).2、Baba T , Mukaida N . Role of macrophage inflammatory protein (MIP)-1α/CCL3 in leukemogenesis[J]. Molecular & Cellular Oncology, 2014, 1(1):-.3、Int. J. Mol. Sci. 2020, 21, 5186

CD20(Cluster of Differentiation 20)是MS4A家族的一员,由297个氨基酸组成的磷蛋白,具有4个跨膜结构域,在B细胞发育中起关键作用。在血液、扁桃体、阑尾、淋巴结和脾脏等多组织表达,正常脑组织和一些免疫细胞上检测不到表达。CD20敲除小鼠与同窝野生型小鼠一样茁壮成长和繁殖,在其出生后第一年未出现任何明显的解剖或形态学异常,或对感染的易感性。CD20没有已知的天然配体,其功能是实现最佳的B细胞免疫应答,特别是针对T细胞非依赖性抗原。CD20表达密度存在B细胞亚群特异性差异,在B细胞淋巴瘤、毛细胞白血病、B细胞慢性淋巴细胞性白血病和黑色素瘤癌干细胞高表达。尽管CD20的功能尚不完全清楚,但它具有动态的细胞活性,是B细胞恶性肿瘤的理想特异性治疗靶标。CD20抗体的6种作用机制[1]CD20抗体CD20抗体是迄今为止最成功的抗肿瘤治疗方案之一,各大药企进行了各种基因工程研究和人源化改造,试图设计和生成强大的CD20抗体。CD20抗体生产的初步成功,推进了单克隆抗体设计方法不断改进。其中4种类型的CD20抗体,包括利妥昔单抗、Ofatumumab、Obinutuzumab和Ublituximab分别于1997年、2010年和2014年在美国和欧洲获批用于治疗慢性淋巴细胞白血病。CD20单克隆抗体设计的不断改进[2]CD20单克隆抗体批准上市时间表及对应症[3]目前,抗 CD20 单抗已经发展到第三代,每一代抗体药物均有各自特有的临床应用价值。01第一代CD20单克隆抗体第一代CD20抗体以利妥昔为代表,针对B细胞上CD20抗原的利妥昔单抗是一种基因工程嵌合鼠/人单克隆抗体,是全球第一款CD20单克隆抗体(mAb)上市药物,上市后迅速成为治疗某些B细胞恶性肿瘤特定类型的标准方案,延续至今。美罗华@利妥昔单抗注射液第一代CD20单抗的疗效很好,甚至对于一直以来的治疗方案产生革命性的影响,但由于其是嵌合鼠/人单克隆抗体,因此使用时会存在一定的排异反应,产生负作用影响治疗效果。为提升治疗效果降低副作用,CD20单抗的药物制备技术不断更新换代,衍生出了第二代CD20单抗。02第二代CD20单克隆抗体第二代抗CD20 mAb将CD20人源化或者全人源改造,降低免疫原性,包括Ofatumumab、Veltuzumab和Ocrelizumab。Ofatumumab (OFA)是一种全人源 I 型抗CD20 IgG1k mAb。Ofatumumab与CD20分子的小和大胞外环(ECL)均结合,在杀伤靶细胞方面比利妥昔单抗更有效。OFA(arzerra)已获批用于治疗氣达拉滨和阿仑单抗治疗失败的复发性或难治性CLL(FA-ref),目前正在开发OFA联合其他药物治疗各种B细胞肿瘤。第二代CD20单克隆抗体[4]与一代CD20抗体相比二代CD20抗体免疫原性降低,减少不良反应,但是二代CD20抗体特异性和抗原结合的亲和力有所下降,因此第三代产品应运而生。03第三代CD20单克隆抗体第三代人源化CD 20 mAb具有Fc工程化改造,以增加其与FcyRIIIa受体的结合亲和力,包括 Ocaratuzumab、PR0131921和Obinutuzumab,均在进行不同适应症的临床开发。Ocaratuzumab(AME-133v)是一种 I 型人源化IgG1 mAb。其与CD20的结合亲和力增加13-20倍,与FcyRIIIa受体低亲和力(F/F和F/V)变体的亲和力增加5-7倍。这些可能是克服利妥昔单抗缓解率较低和缓解持续时间较短的机制。Obinutuzumab(GA101)在2013 年已获得美国 FDA 批准上市,主要适用于未经治疗的慢性淋巴细胞白血病患者以及联合苯达莫司汀二线治疗复发性滤泡性淋巴瘤。第三代CD20单抗[4]虽然CD20单抗的药物制备技术经过多次更新换代,二代三代各具优势,但与一代产品利妥昔单抗相比,在适应症和疗效上并未更具优势,因此目前市场上第一代产品利妥昔单抗依然占据绝对地位。发展前景CD20 单抗是世界上第一个上市的单克隆抗体,具有多年的临床治疗数据,对应不同的适应症以及设计优化,该靶点尚具有巨大的市场潜力。此外基于CD20靶点的研究的人源化单克隆抗体、双特异性抗体、CAR-T疗法、抗体偶联药物(ADC)、与小分子药物联合治疗等研究,均取得良好的效果。国内外有多家企业展开针对CD20靶点的单抗,双抗以及联用药物的研究,超110多个研究管线处于临床前期或已进入临床阶段,超23款药物已上市。今年9月,北京神州细胞自主研发的新型抗CD20单抗——“安平希”瑞帕妥单抗获批上市,是国产首款抗CD20单抗上市药物。国内外部分CD20药物临床研发进展数据来源:Cortellis通过众多临床试验及以往多年研究积累,证实了 CD20 靶点是治疗 B 细胞淋巴瘤的重要靶点,以CD20 为靶点研发出的药物在临床治疗 B 细胞淋巴瘤中发挥了关键作用。百奥动物自主研发的CD20系列人源化小鼠及细胞系,是靶向CD20抗体开发相关药物进行药效评价的优质模型。B-hCD20 mice01基本信息02mRNA表达分析RT-PCR分析CD20基因在野生型小鼠和B-hCD20小鼠中的表达情况。鼠CD20 mRNA仅在野生型(+/+)小鼠脾细胞中可检测到。人CD20 mRNA仅在纯合子B-hCD20(H/H)小鼠中检测到,而在野生型(+/+)小鼠中检测不到。03蛋白表达分析 用流式细胞术分析B-hCD20纯合子小鼠CD20的蛋白表达。采集野生型C57BL/6小鼠和纯合子B-hCD20(H/H)小鼠脾细胞,用种属特异性抗CD20抗体进行流式细胞术分析。结果显示:鼠CD20仅在野生型C57BL/6(+/+)小鼠中检测到;人CD20仅在纯合子B-hCD20(H/H)小鼠中检测到,在C57BL/6(+/+)小鼠中检测不到。B-hCD28/hCD20 mice01基本信息02蛋白表达分析流式细胞术分析B-hCD28/hCD20纯合子小鼠CD28的蛋白表达。采集野生型C57BL/6(+/+)小鼠和纯合子B-hCD28/hCD20(H/H;H/H)小鼠脾细胞,用种属特异性抗CD28抗体进行流式细胞术分析。结果显示:鼠CD28仅在C57BL/6(+/+)小鼠中检测到;人CD28仅在纯合子B-hCD28/hCD20(H/H;H/H)小鼠中检测到,在C57BL/6(+/+)小鼠中检测不到。流式细胞术分析B-hCD28/hCD20纯合子小鼠CD20特异性表达。采集野生型C57BL/6(+/+)小鼠和纯合B-hCD28/hCD20 (H/H;H/H)小鼠脾细胞,用种属特异性抗CD20抗体进行流式细胞术分析。结果显示:鼠CD20仅在C57BL/6(+/+)小鼠中检测到;人CD20仅在纯合子B-hCD28/hCD20(H/H;H/H)小鼠中检测到,在C57BL/6(+/+)小鼠中检测不到。B-hCD3E/hCD20 mice01基本信息02蛋白表达分析流式细胞术分析B-hCD3E/hCD20纯合子小鼠CD3E的蛋白表达。采集野生型C57BL/6(+/+)小鼠和纯合子B-hCD3E/hCD20(H/H;H/H)小鼠脾细胞,用种属特异性抗CD3E抗体进行流式细胞术分析。结果显示:鼠CD3E仅在C57BL/6(+/+)小鼠中检测到;人CD3E仅在纯合子B-hCD3E/hCD20(H/H;H/H)小鼠中检测到,在C57BL/6(+/+)小鼠中检测不到。流式细胞术分析B-hCD3E/hCD20纯合子小鼠CD20的蛋白表达。采集野生型C57BL/6(+/+)小鼠和纯合子B-hCD3E/hCD20(H/H;H/H)小鼠脾细胞,用种属特异性抗CD20抗体进行流式细胞术分析。结果显示:鼠CD20仅在C57BL/6(+/+)小鼠中检测到;人CD20仅在纯合子B-hCD3E/hCD20(H/H;H/H)小鼠中检测到,而在C57BL/6(+/+)小鼠中检测不到。03免疫分型脾脏白细胞亚群的流式分析。分离野生C57BL/6(+/+)小鼠和B-hCD3E/hCD20 (H/H;H/H)小鼠(雌性,n=3,6周龄)的脾细胞并进行白细胞亚群流式分析。A.代表性单活CD45+细胞的流式分析。B.图A的统计学分析。结果显示:纯合子B-hCD3E/hCD20小鼠的T细胞、B细胞、NK细胞、树突状细胞、粒细胞、单核细胞和巨噬细胞的比例与C57BL/6小鼠相似,证明人源化CD3E/CD20不影响脾脏中这些细胞集群总体的发育、分化或分布。数值:平均值 ± SEM脾脏T细胞亚群的流式分析。分离野生型C57BL/6(+/+)小鼠和B-hCD3E/hCD20 (H/H;H/H)小鼠(雌性,n=3,6周龄)的脾细胞并进行T细胞亚群流式分析。A.代表性CD3+ T细胞的流式图分析。B. 图A的统计学分析。结果显示:纯合子B-hCD3E/hCD20小鼠的CD8+ T细胞、CD4+ T细胞和Treg细胞的比例与C57BL/6小鼠相似,证明人源化CD3E/CD20不影响脾脏中这些T细胞集群总体的发育、分化和分布。数值:平均值 ± SEM人源化CD3E/CD20同样也不影响淋巴结、血液中白细胞亚群和T细胞亚群总体的发育、分化和分布。(数据未展示)B-hCD3EDG/hCD20 mice01基本信息02mRNA表达分析RT-PCR分析CD3DG基因在野生型小鼠和B-hCD3EDG/hCD20小鼠中的表达。小鼠Cd3d和Cd3g mRNA仅在野生型(+/+)小鼠胸腺细胞中检测到。人CD3D和CD3G mRNA仅在纯合B-hCD3EDG/hCD20(H/H;H/H)小鼠中检测到,而在野生型(+/+)小鼠中检测不到。03蛋白表达分析流式细胞术分析纯合B-hCD3EDG/hCD20小鼠中CD3E的蛋白表达。采集野生型C57BL/6 (+/+)小鼠和纯合子B-hCD3EDG/hCD20(H/H;H/H)小鼠脾细胞,用种属特异性抗CD3E抗体进行流式细胞术分析。结果显示:鼠CD3E仅在C57BL/6 (+/+)小鼠中检测到;人CD3E仅在纯合子B-hCD3EDG/hCD20 (H/H;H/H)小鼠中检测到,而在C57BL/6 (+/+)小鼠中检测不到。流式细胞术分析B-hCD3EDG/hCD20纯合子小鼠中CD20的蛋白表达。采集野生型C57BL/6 (+/+)小鼠和纯合子B-hCD3EDG/hCD20 (H/H;H/H)小鼠脾细胞,采用种属特异性抗CD20抗体进行流式细胞术分析。结果显示:鼠CD20仅在C57BL/6 (+/+)小鼠中可检测到;人CD20仅在纯合子B-hCD3EDG/hCD20(H/H;H/H)小鼠中检测到,而在C57BL/6 (+/+)小鼠中检测不到。CD20靶点相关模型列表参考文献:1. Payandeh, Z. et al. The applications of anti-CD20 antibodies to treat various B cells disorders. Biomed Pharmacother 109, 2415-2426 (2019).2. Lim, S.H. et al. Anti-CD20 monoclonal antibodies: historical and future perspectives. Haematologica 95, 135-143 (2010).3. Marshall, M.J.E., Stopforth, R.J. & Cragg, M.S. Therapeutic Antibodies: What Have We Learnt from Targeting CD20 and Where Are We Going? Front Immunol 8, 1245 (2017).4. Shundong Cangl, N.M., Kemeng Wang2 and Delong Liu2. Novel CD20 monoclonal antibodies for lymphoma therapy. Cang et al. Journal of Hematology & Oncology 5 (2012).

药物靶点,即药物与人体生物大分子的结合部位,是新药发现的源头。目前全世界在研的抗体类药物靶点有一千多个,如何在其中找到好的药物靶点,开发出一类新药是所有创新药企面临的共同问题。靶点基因的鉴定和验证在药物发现、开发过程中是必不可少且至关重要的一步。在哺乳动物生理学的背景下,基因敲除动物的表型研究已经成为破译基因功能及靶点验证的有力手段。与其他技术相比,基因敲除模型在药物靶点发现方面有自身独特的优势,敲除动物模拟了该靶点被完全抑制时的表型,因而能帮助我们了解该靶点在疾病中的作用,增加我们对人类正常生理过程及疾病发生机制的理解。图片来源 [1]小鼠作为在临床前生物医学研究中常用的模式动物之一,具有诸多方面的优势[2]:小鼠和人类有着绝大部分同源序列,尽管二者在生理和免疫学存在一些差异,但基因组测序发现,小鼠和人类的~30,000个基因中,只有300个(即1%)是两个物种特有的;在一项研究中所有小鼠通常具有相同的遗传背景(即单一近交系或回交到同一个遗传背景)。因此,所有研究对象的基因是相同的,消除了如遗传变异和基因多态性等潜在的复杂因素影响;小鼠繁殖周期短,代际间隔小于3个月,且每窝产仔数量多,可以快速扩繁获得具有统计学意义数量的实验用鼠;在特定的研究中小鼠具有明确的病史,且所有动物可在相同的环境下饲养,排除了其对实验结果可重复性的影响;在一项特定的研究中,所有小鼠采用相同的实验方案,并可进行终点病理学的完整评估,包括检测生理参数和组织病理学分析;小鼠基因组计划已经完成且小鼠基因改造技术成熟,可以构建模拟人类疾病的转基因小鼠模型以了解发病机制及对药物的反应等研究。转基因小鼠模型在生物医学研究中的应用[2]基因敲除小鼠模型除了可以提供表型分析信息外,其在抗体产生和鉴定方面具有两个优势。(1)与野生型小鼠相比,理论上特定基因敲除小鼠产生抗体的效率应该更高,因为敲除小鼠的免疫系统从未接触过免疫蛋白。(2)用于验证抗体特异性,“……严格控制抗体特异性,要求将野生型组织或细胞中的抗体反应性与敲除动物中的抗体反应性进行比较……”。[3] 因此,基因敲除小鼠是开发新型药物或新治疗策略非常有用的实验工具。百奥动物开发了一系列靶点基因敲除(KO)小鼠,可以满足新药开发中“靶点发现”过程的动物模型需求,助力新药研发。扫描下方二维码查看700+基因敲除小鼠资源库,下面以B-IL4ra KO mice(C)为例进行部分数据介绍。B-IL4ra KO mice(C)哮喘小鼠模型构建实验动物:BALB/c,B-IL4ra KO mice(C),4-5周龄,雌性;致敏阶段:第0,7,14天腹腔注射OVA+AI(OH)3;激发阶段:小鼠在第21-25天每天接受2%的OVA雾化30分钟。检测分析(B)哮喘小鼠的支气管肺泡灌洗液(BALF)中免疫细胞浸润分析;(C)血清中IgE的检测;(D)肺组织的HE染色。通过对IL4RA靶点敲除小鼠的哮喘模型数据分析发现:Il4ra基因敲除后,与对照组相比,BALF中浸润的嗜酸性粒细胞数量和比例有显著性下降;血清中检测不到IgE;肺组织中浸润的炎性细胞和粘液分泌减少。这些结果证明IL4RA可以作为哮喘治疗的潜在靶点进行开发。更多小鼠数据信息,欢迎联系我们。参考资料[1] Disease Models & Mechanisms (2016) 9, 101-103. doi:10.1242/dmm.024547[2] Transgenic Res (2012) 21:327–349. doi:10.1007/s11248-011-9537-3[3] Disease Models & Mechanisms (2019) 12, dmm038224.[4] Drug Discovery Today, Volume 17, Supplement, February 2012, Pages S24-S30. https://doi.org/10.1016/j.drudis.2011.09.007
系统性红斑狼疮(Systemic Lupus Erythematosus, SLE)是一种严重的自身免疫性疾病,主要特征为免疫调节功能紊乱,通过自身反应性细胞和抗体可引发炎性细胞因子的过度产生和攻击正常组织(心脏、关节、皮肤、肺、血管、肝脏、肾脏和神经系统)[1]。系统性红斑狼疮发病机制综述[1]遗传、环境、激素、表观遗传和免疫调节因素依次或同时作用于免疫系统。致病因素的作用导致自身抗体、免疫复合物、自身反应性或炎症性T细胞和炎症细胞因子的产生,这些细胞可能启动和放大各种器官的炎症和损伤。受局部因素影响,靶器官可能进一步受损。流行病学数据显示,我国SLE疾病负担较重,预估发病率约为8.57/10万人年,位居全球第四位[2]。其中约90%的SLE患者是女性,其余10%为男性及儿童。研究显示,SLE患者5年生存率从20世纪50年代的50%~60%升高至90年代的超过90%,并在2008年~2016年逐渐趋于稳定。SLE已由既往的急性、高致死性疾病转为慢性、可控性疾病[3]。SLE传统治疗药物包括糖皮质激素、抗疟药、免疫抑制剂、富马酸酯、间充质干细胞、小剂量白细胞介素2(IL2)、生物制剂等。目前,获批上市用于治疗SLE的靶向生物药仅3种。BioMice百奥动物自主开发了一系列稳定的靶点人源化小鼠模型,能够在不同方面模拟SLE发生发展过程,为临床前开发、评价更有效的SLE治疗药物提供了相应的疾病模型,加速了相关药物的研发进程。部分模型数据展示如下:>>>B-hTWEAK/hAPRIL mice肿瘤中的TWEAK/Fn14信号通路[4]。将TWEAK与Fn14结合导致Ect2 GEF激活Cdc42,随后由Trio GEF激活Rac1,促进胶质瘤细胞的迁移和侵袭。TWEAK/Fn14信号同时激活经典和替代NF-κB通路,诱导细胞因子、趋化因子、粘附分子的产生。TWEAK/Fn14相互作用激活TRAF2信号通路。然后,Fn14招募一个cIAP1 TRAF2复合体。cIAP1使细胞对TNF-α敏感。TNF-α通过与TNFR1和TNFR2两种受体结合产生不同的效应。TNFR1可诱导细胞死亡,TNFR2可促进细胞增殖。BAFF和APRIL受体相互作用[5]。B细胞刺激分子BAFF和APRIL是维持B细胞和体液免疫的关键因素。BAFF和APRIL可以裂解成三聚体可溶性细胞因子,也可以组装成异构体。BAFF结合它的三个受体具有不同的亲和力:它与BAFF-R结合最强,其次是TACI,与BCMA结合较弱。APRIL是BCMA的首选配体,其次是TACI。APRIL还在细胞外基质和肿瘤或其他细胞表面与HSPG结合,触发APRIL多聚化并通过TACI实现信号传递。BAFF通过BAFF-R信号通路使用NF-κB和PI3K通路,而APRIL信号通过NF-κB通路与BCMA和TAC结合。B细胞蛋白表达分析>>>纯合B-hTWEAK/hAPRIL小鼠中种属特异性APRIL表达的流式细胞术分析。采集野生型(WT)小鼠和纯合B-hTWEAK/hAPRIL小鼠脾细胞,用抗APRIL抗体流式细胞术分析。由于抗体的交叉反应性,在WT小鼠和纯合B-hTWEAK/hAPRIL小鼠中检测到APRIL。>>>B-hBAFF mice人B细胞激活因子和增殖诱导配体抑制剂的作用机制[6]。B细胞激活因子(BAFF)和增殖诱导配体(APRIL)与三种受体B细胞激活因子受体(BAFF-R)、跨膜激活剂和钙调节剂和亲环配体相互作用剂(TACI)和B细胞成熟抗原(BCMA)结合不同。选择性BAFF抑制剂阻断可溶性BAFF或可溶性和膜性BAFF与其受体之间的相互作用,使APRIL功能完整,而BAFF/APRIL双抑制剂atacicept (TACI-Ig)阻断BAFF和APRIL与所有三种受体的相互作用。BAFF抑制会消耗B细胞并改变自身反应性B细胞的选择,并可能对T细胞和树突状细胞(DC)有直接或间接的影响。蛋白表达分析>>>用ELISA法分析野生型(WT)小鼠和B-hBAFF小鼠中BAFF的表达。采集WT小鼠和杂合B-hBAFF小鼠血清,采用种属特异性BAFF ELISA试剂盒进行ELISA分析。在WT小鼠和杂合B-hBAFF小鼠(H/+)中检测到小鼠BAFF。人BAFF仅在杂合B-hBAFF小鼠中检测到。ND:检测不到。>>>B-hBAFFR mice蛋白表达分析>>>纯合B-hBAFFR小鼠中种属特异性BAFFR表达的流式细胞仪分析。取WT和纯合B-hBAFFR小鼠脾细胞,用种属特异性抗BAFFR抗体流式细胞术分析。小鼠BAFFR在WT小鼠中检测到。人BAFFR仅在纯合B-hBAFFR中检测到,而在WT小鼠中检测不到。>>>B-hBAFF/hBAFFR mice蛋白表达分析>>>流式细胞术分析纯合B-hBAFF/hBAFFR小鼠中种属特异性BAFFR的表达。从野生型(WT)小鼠和纯合B-hBAFF/hBAFFR小鼠采集脾脏、淋巴结和血液中的B细胞,用种属特异性抗BAFFR抗体流式细胞仪分析。在WT小鼠中检测到小鼠BAFFR。人BAFFR仅在纯合B-hBAFF/hBAFFR小鼠检测到,而WT小鼠(+/+)则没有。>>>B-hC1Q miceC1q在免疫稳态中具有基本的抑制作用[7]。a.在描述良好的凋亡细胞清除通路中,C1q与吞噬细胞上的各种gC1q和C1q尾部受体相互作用,导致细胞因子产生/炎症反应的调节。C1q胶原蛋白尾部与LAIR-1之间的相互作用可阻止pDCs和单核细胞产生I型ifn和炎症因子,并在稳态或炎症状态下抑制DC分化和激活。PDC浆细胞样树突状细胞,Mono单核细胞,moDC单核细胞来源的树突状细胞。蛋白表达分析>>>ELISA法检测C1Q在野生型C57BL/6小鼠和纯合B-hC1Q小鼠中的表达。采集野生型C57BL/6小鼠和纯合B-hC1Q小鼠血清。小鼠C1Q仅在野生型小鼠中检测到。C1Q在野生型C57BL/6小鼠和纯合B-hC1Q小鼠中均可检测到。因此,推测该抗人C1Q抗体在人鼠之间具有交叉反应性。>>>B-hCLEC4C miceBDCA-2靶点全名为血液树突状细胞抗原2,也叫CLEC4C或CD303。CD303与FcRγ-链相关,在BCR样信号体中发出信号。触发CD303导致Syk的激活,SLP65的募集和PLCγ2的活性。在pDC中,PLCγ2-PKC通路可能在CD303触发后对NF-kB通路起负调控作用,并抑制IFN-I基因。越来越多的证据表明,pDC作为IFN-I的主要生产者可能在SLE的发病机制中发挥作用。pDC衍生的I型IFN被认为在SLE的致病性中发挥重要作用。血液树突状细胞抗原(BDCA2)是一种受体,特异性表达于人类和非人类灵长类动物的pDCs上,抑制TLR7和TLR9介导的I型IFN[8,9]。蛋白表达分析>>>流式细胞术分析B-hCLEC4C小鼠中种属特异性CLEC4C表达。采集野生型C57BL/6小鼠、杂合B-hCLEC4C小鼠和纯合B-hCLEC4C小鼠脾细胞,用抗CLEC4C抗体进行流式细胞术分析。人类CLEC4C仅在B-hCLEC4C小鼠的浆细胞样树突状细胞(pDCs)中检测到,而在野生型小鼠中检测不到。在B-CLEC4C小鼠和野生型小鼠的T细胞、B细胞和NK细胞中均未检测到人CLEC4C。参考资料:[1] Tsokos GC. Systemic lupus erythematosus. N Engl J Med. 2011, 365(22):2110-21.[2] Tian J, Zhang D, Yao X, et al. Global epidemiology of systemic lupus erythematosus: a comprehensive systematic analysis and modelling study. Ann Rheum Dis. 2022 Oct 14:ard-2022-223035.[3]曾小峰,陈耀龙.2020中国系统性红斑狼疮诊疗指南[J].中华内科杂志,2020(3):172-185.[4] Hu G, Zeng W, Xia Y. TWEAK/Fn14 signaling in tumors[J]. Tumor Biology, 2017, 39(6): 1010428317714624.[5]Samy E, Wax S, Huard B, et al. Targeting BAFF and APRIL in systemic lupus erythematosus and other antibody-associated diseases[J]. International reviews of immunology, 2017, 36(1): 3-19.[6]Boneparth A , Davidson A . B-cell activating factor targeted therapy and lupus[J]. Arthritis Research & Therapy, 2012, 14(4 Supplement):S2-S2.[7]Son M, Diamond B, Santiago-Schwarz F. Fundamental role of C1q in autoimmunity and inflammation. Immunol Res. 2015;63(1-3):101-106. doi:10.1007/s12026-015-8705-6[8]Röck J, Schneider E, Grün JR, et al. CD303 (BDCA-2) signals in plasmacytoid dendritic cells via a BCR-like signalosome involving Syk, Slp65 and PLCgamma2. Eur J Immunol. 2007;37(12):3564-3575. doi:10.1002/eji.200737711[9]Pellerin A, Otero K, Czerkowicz JM, et al. Anti-BDCA2 monoclonal antibody inhibits plasmacytoid dendritic cell activation through Fc-dependent and Fc-independent mechanisms. EMBO Mol Med. 2015;7(4):464-476. doi:10.15252/emmm.201404719

免疫系统人源化小鼠是研究人造血和免疫功能的有力工具。在B-NDG小鼠或B-NDG hIL15小鼠中移植人CD34+ HSCs后,可以很好地重建人淋系细胞包括T细胞、B细胞和NK细胞,用于各类淋系细胞的作用机制研究及药效评价。但在这些人源化小鼠模型中,人髓系细胞包括单核/巨噬细胞、树突状细胞、粒细胞和红细胞等重建不足,主要原因是促进各类髓系细胞重建的细胞因子在人和鼠之间的交叉反应不足。已有的数据表明:人细胞因子IL3、人粒细胞-巨噬细胞集落刺激因子 (GM-CSF) 和人巨噬细胞集落刺激因子(M-CSF) 在小鼠中的表达可以促进树突状细胞、单核/巨噬细胞的发育与分化;人血小板生成素(THPO)能促进功能性人造血干细胞维持其多向分化潜能并促进其在小鼠骨髓中的长期植入。成熟血细胞的寿命是有限的,需要不断更新。造血从骨髓中一小部分多能造血干细胞的增殖和分化开始。造血程序异常会破坏体内平衡,并促使骨髓、血液或外周淋巴器官中中间祖细胞或成熟细胞的积累,从而导致各种恶性肿瘤。图1.造血作用机制[1]IL31981年Ihle等发现ConA刺激小鼠脾细胞的培养上清中含有一种因子,能提高裸鼠脾脏淋巴细胞成熟标志物20-α-羟固醇氢酸阳性率。这个因子被称为白细胞介素3 (IL-3);因其功能可刺激多功能干细胞和多种祖细胞的增殖与分化,又称为多重集落刺激因子(multi-CSF)。IL-3可刺激皮肤上皮细胞、CD4-CD8-TCRαβ细胞、肥大细胞、嗜碱性粒细胞增殖,并阻止肥大细胞发生程序性细胞死亡等。GM-CSFGM-CSF,又称CSF-2,是一种分泌型细胞因子。GM-CSF有广泛的生理功能,主要参与髓细胞的生成和分化成熟、促进M1型巨噬细胞极化、激活中性粒细胞、参与血管形成、肿瘤进展、炎症发生等。GM-CSF的异常表达会导致过度的炎症、疼痛和组织损伤,并促进其他炎症细胞因子的产生。新冠病毒感染通常会引起淋巴细胞减少和炎症性细胞因子的释放,炎症风暴被认为是患者死亡的重要原因之一,一些医院正在尝试通过IL-6单抗应对新冠病毒引发的炎症风暴。但中科院魏海明教授等人的发现,GM-CSF在新冠病毒感染重症患者中或许更重要。M-CSF巨噬细胞集落刺激因子为M-CSF,又称为集落刺激因子-1(CSF-1)。最初发现于血清、尿或其它体液中,能刺激骨髓造血细胞巨噬细胞集落的形成。M-CSF对破骨细胞的分化与调节,单核细胞的增殖,分化及维持活性有重要作用。THPO促血小板生成素(THPO)能够促进巨核细胞形成和分化,并向外周血释放成熟的血小板,而肿瘤细胞分泌的一种细胞因子白细胞介素-6(IL-6)能够刺激肝脏分泌更多的THPO,因而提高癌症病患血浆血小板浓度。图2.肿瘤细胞利用血小板促进自身的侵袭和迁移[2]百奥动物将重度免疫缺陷小鼠B-NDG mice的IL3、GM-CSF、M-CSF、THPO基因的全长序列分别替换为相应人基因的全长编码序列,并通过二次基因编辑及相互交配的方式获得4种细胞因子人源化的的重度免疫缺陷小鼠,即B-NDG MGMT3 mice。移植人CD34+HSCs后,与B-NDG小鼠相比,B-NDG MGMT3 mice中各类人髓系细胞的重建明显增强,包括单核/巨噬细胞、树突状细胞;人淋系细胞包括CD4+ T细胞、Tregs和NK细胞的重建水平也有增加。B-NDG MGMT3 mice可用于研究造血系统的发育与分化机制、研究肿瘤、自免、感染与代谢等免疫相关疾病的致病机制与药效评价。B-NDG MGMT3 mice蛋白表达分析ELISA法检测野生型B-NDG小鼠和纯合B-NDG MGMT3小鼠中种属特异性GM-CSF、CSF1和THPO的表达。采集LPS刺激小鼠血清,进行ELISA分析(n=3)。小鼠GM-CSF、CSF1和THPO仅在B-NDG小鼠(+/+)中检测到,而在B-NDG MGMT3小鼠(H/H)中检测不到。人GM-CSF、CSF1和THPO仅在B-NDG MGMT3小鼠中检测到。由于IL3主要在活化的T细胞中表达,而B-NDG背景小鼠中没有成熟的T细胞,因此在两种小鼠中均未检测到小鼠或人IL3。利用B-NDG MGMT3 mice移植人CD34+HSCs重建人淋系和髓系细胞将人CD34+HSC(3E4)分别经面部颞静脉植入出生24-72小时的B-NDG小鼠和B-NDG MGMT3小鼠(雌雄均有,n=15)。B-NDG小鼠提前经1.0 gy辐照预处理;B-NDG MGMT3小鼠不做辐照预处理。结果显示:与B-NDG小鼠相比,B-NDG MGMT3小鼠未辐照,移植人HSCs 24周后,其生存率从移植18周开始降低,24周结实验时达到42.85%;但小鼠体重明显较大,重建过程中稳定增长。B-NDG MGMT3小鼠中重建的所有淋系及髓系人免疫细胞数量从重建12周开始显著高于在B-NDG小鼠中的重建水平。B-NDG®miceB-NDG小鼠是百奥赛图自主研发,通过在NOD scid背景上敲除IL2rg基因构建而成的重度免疫缺陷小鼠。该小鼠缺乏成熟的T、B和功能性的NK细胞,并表现出细胞因子信号转导功能缺失,其重度免疫缺陷表型使得该小鼠能够被用来进行包括人免疫细胞在内的异种细胞移植实验。该模型广泛应用于肿瘤学、肿瘤免疫学、传染病和干细胞生物学等领域的研究和药物发现。免疫缺陷系列大小鼠列表参考文献[1] Springuel L, Renauld JC, Knoops L. JAK kinase targeting in hematologic malignancies: a sinuous pathway from identification of genetic alterations towards clinical indications. Haematologica. 2015 Oct;100(10):1240-53. doi: 10.3324/haematol.2015.132142. PMID: 26432382; PMCID: PMC4591756.[2] Monika Haemmerle,Rebecca L. Stone,David G. Menter. The Platelet Lifeline to Cancer: Challenges and Opportunities. Cancer cell. VOLUME 33, ISSUE 6, P965-983, JUNE 11, 201
肿瘤相关髓系细胞(tumor-associated myeloid cells, TAMC)是肿瘤微环境(TME)中重要的组成部分,具有异质性,在肿瘤微环境中可以发挥不同,甚至是相反的作用,如免疫抑制或免疫刺激。靶向TAMCs作为单一疗法或与化疗、免疫疗法联合应用的研究正在火热进行中。深入研究TAMCs在肿瘤中的功能和作用机制将有助于发现新的治疗方法。研究最多的肿瘤相关髓系细胞包括单核细胞、肿瘤相关巨噬细胞(TAMs)、树突状细胞(DC)、癌症相关循环中性粒细胞、肿瘤相关中性粒细胞(TANs)和骨髓来源抑制细胞(MDSCs)。TAMC的主要功能是调节淋巴细胞行为进而形成免疫刺激或免疫抑制性TME,从而抑制或促进包括肿瘤细胞的恶性克隆进化、生长、存活、侵袭、播散和转移、血管生成在内的各个肿瘤发展阶段。单核细胞单核细胞是一组异质性的单核吞噬细胞,分为经典型单核细胞、中间型单核细胞和非经典型单核细胞,在炎症期间循环外周血中发挥先天免疫功能。肿瘤发展的不同阶段,不同的单核细胞亚群表现出不同甚至相反的作用。从机制上讲,中间型单核细胞经肿瘤细胞刺激后,促炎细胞因子TNF- α和白细胞介素12(IL-12)的产生增加,而抗炎细胞因子白细胞介素10(IL-10)的产生减少,并对肿瘤细胞产生直接的细胞毒性,促进肿瘤细胞凋亡。经典型单核细胞产生VEGF,促进肿瘤细胞外渗,导致转移。相比之下,非经典型单核细胞在吞噬肿瘤细胞衍生微粒后被激活,从而减少肿瘤细胞的转移。肿瘤中单核细胞的功能肿瘤相关巨噬细胞(TAMs)巨噬细胞作为单核吞噬细胞系统,在组织稳态和炎症中起着关键作用。巨噬细胞分为两个主要亚群,M1和M2巨噬细胞,在功能上是异质的。M1巨噬细胞是对抗微生物感染的第一道防线,具有很强的抗原呈递能力诱导强烈的Th1反应。在脂多糖(LPS)、IFN-γ和粒细胞-巨噬细胞刺激因子(GM-CSF)的作用下,M1巨噬细胞经历经典激活,并优先分泌抗菌分子和促炎细胞因子。M2巨噬细胞在限制免疫反应、诱导血管生成和组织修复方面起着关键作用。在白细胞介素4(IL-4)、白细胞介素13(IL- 13)、IL-10和CSF-1的作用下,M2巨噬细胞发生选择性激活并优先分泌抗炎细胞因子。TAMs在肿瘤中的作用树突状细胞(DC)DC是最有效的抗原呈递细胞(APCs),连接先天免疫和适应性免疫,在生理条件下具有表型和功能异质性。在对微生物感染的反应中,细胞外微生物蛋白通常被成熟DC吞噬或内吞,并通过MHC II分子呈递给CD4+ T细胞。相比之下,胞质微生物蛋白通常通过MHC I分子呈现给CD8+ T细胞。TME浸润的DC包括不同发育阶段的树突状细胞亚群,这些肿瘤相关树突状细胞根据不同的细胞亚群和肿瘤分期,发挥免疫刺激或免疫抑制作用。DC在肿瘤中的作用粒细胞粒细胞可大致分为癌症相关循环中性粒细胞、肿瘤相关中性粒细胞(TANs)以及其他中性粒细胞。癌症相关循环中性粒细胞这些与癌症相关的循环中性粒细胞由功能异质性亚群组成,是循环外周血的多形核吞噬细胞,具有对抗微生物病原体的先天免疫功能。在癌症患者中,特别是在晚期和转移后,循环中性粒细胞数量增加,高中性粒细胞与淋巴细胞比率(NLR)与侵袭性癌症相关。肿瘤相关中性粒细胞(TANs)研究表明TME中的TANs可促进肿瘤细胞增殖、外渗和迁移,在肿瘤发展和转移中起着关键作用。TANs可释放弹性蛋白酶等颗粒内容物,促进肿瘤增殖和侵袭细胞;分泌IL-1β和MPPs促进肿瘤细胞外渗到转移前壁龛,从而促进癌细胞的扩散;激活TLR通路,促进肿瘤细胞的迁移、黏附、侵袭和转移。除促肿瘤作用外,TANs还被证明通过产生ROS和TRAIL介导肿瘤细胞的细胞毒性。研究也表明TANs与TME中的淋巴细胞相互作用并调节其功能。其他粒细胞除了嗜中性粒细胞(neutrophilic),其他类型的粒细胞,如嗜酸性粒细胞(eosinophils)和嗜碱性粒细胞(basophils),对肿瘤也有影响。研究表明嗜酸性粒细胞具有抗肿瘤活性,对各种癌细胞都表现出直接的或间接的细胞毒性,从而抑制肿瘤生长。中性粒细胞在肿瘤中的作用骨髓来源抑制细胞(MDSCs)MDSCs是一组异质性的髓系祖细胞和处于不同发育阶段的未成熟髓系细胞,它们在血管中循环。在感染时,MDSCs迅速扩张并分化为粒细胞、单核细胞、巨噬细胞和树突状细胞,在调节免疫反应和组织修复中发挥重要作用。肿瘤相关的MDSC由两个主要亚群体组成,粒细胞骨髓来源抑制细胞(GrMDSCs)和单核细胞骨髓来源抑制细胞(MoMDSCs)。GrMDSCs在表型和形态上与中性粒细胞相似,MoMDSCs在表型和形态上与单核细胞相似,研究表明GrMDSCs和MoMDSCs均参与T细胞介导的免疫抑制。MDSCs及其亚群在肿瘤中的作用髓系细胞在肿瘤免疫治疗的研究中的作用是毋庸置疑的,调节这些髓系细胞的发育、成熟和功能有助于发现新的肿瘤免疫治疗策略。然而,由于各种可互换亚群的复杂性和可塑性,这些髓系细胞执行重叠或相反的功能,目前控制TAMCs行为的分子机制在很大程度上尚不清楚。为进一步阐明髓系细胞各亚群在不同癌症中的功能,并确定其促肿瘤和抗肿瘤活性相关的分子机制,百奥动物自主研发一些列髓系靶点人源化小鼠,助力深入了解髓系细胞的复杂性,并设计新的肿瘤靶向治疗方法。B-hTREM2 mice(C) 基本信息体内药效抗人TREM2抗体在B-hTREM2(C)小鼠中的抗肿瘤活性。纯合子B-hTREM2(C)小鼠(雌性,10周龄,n=6)小鼠皮下接种小鼠乳腺癌EMT-6细胞。当肿瘤体积达到约50-80 mm3时,对小鼠进行分组,然后使用抗人TREM2抗体进行治疗。如图A所示,抗人TREM2抗体在B-hTREM2(C)小鼠中有效地控制肿瘤生长,证实B-hTREM2(C)小鼠模型是抗人TREM2抗体临床前体内药效评估的优质模型。数值以平均值±SEM表示。B-hCD36 mice基本信息体内药效抗小鼠PD-1抗体和抗人CD36抗体联合治疗在B-hCD36小鼠中的抗肿瘤活性。纯合子B-hCD36小鼠(雌性,7周龄,n=5)小鼠皮下接种小鼠结肠癌MC38细胞。当肿瘤体积达到约60 mm3时,对小鼠进行分组,然后使用抗体进行治疗。如图A所示,抗体组合在B-hCD36小鼠中有效地控制肿瘤生长,证实B-hCD36小鼠模式是抗CD36抗体临床前体内药效评估的优质模型。数据来自合作者,数值以平均值±SEM表示。B-hPD-1/hPD-L1/hVSIR mice基本信息蛋白表达分析通过流式细胞术分析野生型C57BL/6小鼠和纯合子B-hPD-1/hPD-L1/hVSIR小鼠中种属特异性PD-1和PD-L1的表达。用抗CD3ε抗体体内刺激野生型C57BL/6小鼠和纯合子B-hPD-1/hPD-L1/hVSIR小鼠,收集脾细胞并进行流式分析。结果显示:小鼠PD-1和PD-L1在野生型C57BL/6小鼠中检测到。人PD-1和PD-L1只在纯合子B-hPD-1/hPD-L1/hVSIR小鼠中检测到,而在野生型C57BL/6小鼠中检测不到。通过流式细胞术分析野生型C57BL/6小鼠和纯合子B-hPD-1/hPD-L1/hVSIR小鼠中种属特异性VSIR的表达。取野生型C57BL/6小鼠和纯合子B-hPD-1/hPD-L1/hVSIR小鼠脾细胞进行流式分析。结果显示:小鼠VSIR仅在野生型C57BL/6小鼠中检测到。人VSIR只在纯合子B-hPD-1/hPD-L1/hVSIR小鼠中检测到,而在野生型C57BL/6小鼠中检测不到。髓系靶点人源化小鼠列表参考资料:1. Aixia Dou, Jing Fan. Heterogeneous Myeloid Cells in Tumors. Cancers (Basel). 2021 Aug; 13(15): 3772.2. Sijin Cheng, Ziyi Li, et al. A pan-cancer single-cell transcriptional atlas of tumor infiltrating myeloid cells. Cell. 2021Feb; 184, 792–809.

随着全球人口不断增长与老龄化的加剧,恶性肿瘤已成为威胁人类健康的主要因素,其疾病死亡率一直高居首位。近年来,免疫治疗在抗肿瘤治疗中的作用备受关注,基于免疫治疗的新药开发和标志物探索,成为了当前肿瘤研究的热点,这也对临床前研究中动物模型的建立和使用提出了更高的要求。21世纪以后,人类的生物医学研究主要受限于生物体的复杂性,这使得临床前利用模式动物对抗肿瘤药物的研究评价显得尤为重要。但由于人与小鼠的物种差异和免疫排斥特性,简单的基因改造手段无法在小鼠身上创造出一个和人类免疫系统相似的免疫环境。如小鼠同种异体移植瘤模型、基因工程小鼠、人源性细胞系移植瘤模型、人源性肿瘤组织移植瘤模型等动物模型均缺乏人免疫系统及肿瘤免疫微环境,极大地限制了免疫机制及免疫治疗的转化研究。因此,既可以模拟人肿瘤特征又同时存在“人源化”免疫系统的免疫重建小鼠模型就成为了免疫肿瘤研发中的优质模型。目前应用于临床前的免疫系统人源化小鼠模型主要有三类,一类是将成熟的人外周血单个核细胞(hPBMC)通过腹腔或者尾静脉注入免疫缺陷小鼠体内重建人的免疫系统,即hPBMC型;另一类是将人CD34+造血干细胞(HSC)通过腹腔或者尾静脉注入免疫缺陷小鼠,同样实现了人类免疫系统的重建,即HSC(CD34+)型;还有一类是移植胎儿胸腺和胎肝到经过辐照的重度免疫缺陷小鼠肾包膜下,同时接种人的骨髓造血干细胞进行免疫重建,即BLT模型。本文将重点介绍HSC免疫系统重建的小鼠模型。三种方式免疫重建小鼠[1]HSC具有高度自我更新和多向分化潜能,是各种免疫细胞的共同祖先。HSC的分化依赖于骨髓和胸腺造血微环境,部分分裂增殖以维持数量相对恒定,部分增殖分化成表面标志为CD34+/CD38+的定向干细胞,包括淋巴系祖细胞(CLP)和髓系祖细胞(CMP)。CLP继续分化为T细胞、B细胞和NK细胞。CMP进一步分化为单核-巨噬细胞、中性粒细胞、嗜酸性粒细胞、嗜碱性粒细胞、肥大细胞、红细胞和血小板。HSC发育为成熟免疫细胞的每一阶段均需多种细胞因子的参与。造血干细胞的发育[2]免疫重建模型构建的第一步是要构建免疫缺陷小鼠模型。我们使用的B-NDG小鼠为百奥赛图自主研发的,缺乏成熟的T、B、NK细胞的重度免疫缺陷小鼠模型,是目前国际公认的免疫缺陷程度高、适合人源细胞或组织移植的工具小鼠。第二步即是在B-NDG及B-NDG衍生小鼠的基础上移植入人的免疫细胞、造血干细胞等,构建免疫系统重建小鼠,以能够更好的模拟人的免疫系统,进行免疫学研究和免疫药物评价。免疫重建小鼠模型可用于研究肿瘤在肿瘤微环境中的生长情况以及肿瘤细胞与免疫细胞之间的相互作用。具体可应用于血液疾病、肿瘤免疫、造血和免疫学、人类疾病感染模型,以及免疫抑制剂、双特异性抗体等药物药效评价、ADCC效应功能等的研究。那我们是如何使用B-NDG小鼠建立HSC免疫重建模型,构建后的模型数据又是如何呢?将1.5×105 CD34+ HSC 细胞接种到辐射后的B-NDG免疫缺陷小鼠体内,会使HSC发育成T、B、NK细胞等,从而建立了人的固有免疫系统和淋巴细胞。HSC免疫重建模型相对来说,发生移植物抗宿主病(GvHD)的时间较晚,小鼠的生存期较长。那么拿到HSC(CD34+)型的模式小鼠又能做些什么呢?下面我们来看一组数据,如图所示,在B-NDG小鼠上进行CD34+免疫重建,静脉注射5×105 Raji-Fluc细胞,5天后再给小鼠静脉注射人源PD-1抗体,2天后可见抗体对肿瘤细胞有明显的抑制作用。说明HSC(CD34+)重建的B-NDG小鼠是CDX药效试验的有效模型。此外,我们也可以利用此模型进行双特异性抗体药效评价或者免疫检查点药物和其他药物联用,比如PD-L1抗体与CD47抗体联合用药,这不仅能达到验证人的抗体药效作用,同时由于使用的是人源细胞系或PDX样本,且免疫系统环境高度人源化,实验结果具有更高的可靠性和参考价值。但这种重建方法也存在一些局限性,例如小鼠体内缺乏人的细胞因子,人类干细胞在小鼠体内发育受限等。于是研究人员也尝试通过基因工程敲入人的细胞因子基因来代替小鼠的基因,可以使组织、细胞中人类细胞因子适当表达[3]。huHSC-B-NDG hIL15 miceIL15(interleukin 15)是一种编码白细胞介素家族蛋白的多效细胞因子,在先天性和适应性细胞稳态以及外周免疫功能中发挥重要作用。IL15支持先天淋巴样细胞的发育,通过对IL15转基因小鼠和IL15敲除小鼠的研究表明,IL15在NK细胞、自然杀伤性T细胞(NKT)和记忆性CD8+T细胞的发育中起着至关重要的作用。移植人造血干细胞(HSCs)的小鼠模型表明,人IL15是移植后人NK细胞发育所必需的细胞因子。人NK细胞发育示意图[4]百奥动物开发了B-NDG hIL15小鼠,将人IL15基因的编码序列(CDS)插入小鼠IL15基因的5’UTR后,使小鼠表达人的IL15,但不表达鼠的IL15。该小鼠具有B-NDG小鼠背景,同时表达人IL15,将人HSCs移植到B-NDG hIL15小鼠,不管是成体鼠还是新生鼠,与B-NDG小鼠相比,人NK细胞的重建率都有明显提高。B-NDG hIL15小鼠可以用来研究人NK细胞的发育与功能、评价依赖NK细胞发挥作用特别是具有ADCC功能的抗体药效的有力工具。在B-NDG hIL15小鼠中移植CD34+ HSCs增强人NK细胞的重建(成体鼠)雌性6周龄的B-NDG小鼠(n=17)和B-NDG hIL15小鼠(n=19)分别经1.6Gy照射后,通过尾静脉注射人HSCs (1.5E5),在不同的时间点取外周血检测各类型人免疫细胞的重建水平。结果显示:与B-NDG小鼠相比,B-NDG hIL15小鼠中重建的人NK细胞比例显著性增高,从重建2周到14周都能比较稳定地维持较高比例。利用人HSCs重建人免疫系统的B-NDG hIL15小鼠肿瘤模型能有效验证抗人CLDN18.2抗体的药效(成体鼠)5周龄的B-NDG hIL15小鼠经1.2 Gy照射后,通过尾静脉注射人HSCs (1.5E5), 6周后皮下注射人CLDN18.2过表达的人肺癌B-hCLDN18.2 A549细胞(1E7),7天开始腹腔注射抗人CLDN18.2抗体(zolbetuximab,内部合成),每周取外周血检测人NK细胞和T细胞的重建水平,实验终点时取肿瘤组织检测浸润的人NK细胞和T细胞水平。结果显示:在重建人免疫系统的B-NDG hIL15小鼠肺癌模型中,抗人CLDN18.2抗体能有效抑制肿瘤的生长,肿瘤抑制率是36.2%;外周血及肿瘤组织中都能检测到较高水平的人NK细胞;外周血及肿瘤组织中也能检测到人T细胞,但人T细胞的重建较人NK细胞重建较慢,在外周血中的人T细胞水平维持逐渐升高的趋势。若想减少CD34+ HSC细胞数量的使用,可以使用新生鼠进行免疫重建实验。新生鼠免疫重建实验新生鼠与成体鼠HSC免疫重建对比使用新生鼠移植人CD34+ HSCs重建人免疫系统的方法与特点在新生的B-NDG hIL15小鼠中移植人CD34+ HSCs能重建功能性人NK细胞出生后24-48h的B-NDG小鼠(n=10)和B-NDG hIL15小鼠(n=10)分别经1.0、0.9 Gy辐射后,通过颞静脉注射人CD34+ HSCs (3E4),在不同的时间点取外周血检测各类型人免疫细胞的重建水平。结果显示:与B-NDG小鼠相比,B-NDG hIL15小鼠中重建的人NK细胞比例显著性增高,从重建6周到18周都能比较稳定地维持较高比例。此外,通过将不同种类的肿瘤细胞系移植入免疫重建的小鼠的体内构建的CDX模型,我们可以最大限度地利用小鼠模型模拟人体内免疫系统的响应情况,为研究肿瘤在人体内的生长状态提供了最优模型。百奥动物开发了一系列免疫系统重建的小鼠模型,huHSC-B-NDG hIL15 mice目前保持大量稳定现货供应,欲购从速!百奥赛图自主开发的B-NDG系列小鼠,其成熟的T、B和NK细胞全部缺失,是目前公认的免疫缺陷程度高、非常适合人源细胞或组织移植的工具小鼠,在此基础上百奥动物开发了一系列二代鼠,详情可访问百奥动物官网了解,如有需要请随时与我们联系!参考文献:[1] De La Rochere P, etc. Humanized Mice for the Study of Immuno-Oncology. Trends Immunol. 2018 Sep;39(9):748-763.[2] Reya T, etc. Stem cells, cancer, and cancer stem cells. Nature. 2001 Nov 1;414(6859):105-11. [3] Rongvaux A, etc. Development and function of human innate immune cells in a humanized mouse model. Nat Biotechnol. 2014 Apr;32(4):364-72. [4] Fehniger, etc. Interleukin 15: biology and relevance to human disease. Blood 97, 14-32.

肿瘤治疗药物是全球最大和增长最快的药物细分市场,近年来,疫苗开发也已经成为癌症这一重大疾病新治疗选择的一个热门研究方向。相比全球疫苗市场,全球肿瘤疫苗市场规模增长更快,2019年为46亿美元,预计到2024年将达到101亿美元,复合年增长率为17.28%。01肿瘤疫苗简介疫苗是指含有抗原、能够诱导人体产生特异性主动免疫的制剂,它可以保护机体免受感染源、毒素,以及感染源引起的抗原性物质的损伤。肿瘤疫苗作为肿瘤治疗的新方向,已成为当今肿瘤研究的热点话题。肿瘤疫苗(又称癌症疫苗)通常是含有肿瘤特异性抗原(TSA)或肿瘤相关抗原(TAA)的肿瘤细胞或碎片或片段,可分为肿瘤预防性疫苗(如HPV疫苗)和肿瘤治疗性疫苗(如新抗原mRNA疫苗),主要由肿瘤抗原、制剂、免疫佐剂和递送载体4个关键成分组成。进入人体后,癌症疫苗可激活患者自身免疫系统,诱发特异性免疫反应,克服免疫抑制状态,提高对特定肿瘤的抵抗,是一种主动免疫治疗方法。对于已经发生癌症的患者来说,治疗性疫苗或可延长他们的生存期,成为救命稻草。肿瘤疫苗作为一种新兴的治疗手段,究竟“优秀”在何处呢?肿瘤疫苗组成因素[1]首先,治疗性肿瘤疫苗可通过主动免疫方式诱导全身性的特异性抗瘤效应,与手术治疗、放射治疗相比,作用范围更广泛,特别适用于多发病灶性肿瘤、广泛转移性瘤或非实体性肿瘤(如白血病);其次,肿瘤疫苗可调动机体自身的力量达到抗肿瘤作用,与放射治疗、化学药物治疗相比不良反应小、特异性高。尽管各种治疗性肿瘤疫苗的设计思想各不相同,诱导免疫应答的方式不同,抗瘤效应各有特点,但不论采用何种方法制备,都是围绕如何提高肿瘤抗原的免疫原性以及如何打破肿瘤免疫耐受这一最关键的核心问题展开的。一种理想的肿瘤疫苗不但能够诱导主动性免疫,刺激荷瘤宿主产生有效的免疫应答,同时还应当是安全的和无不良反应的;不仅能特异性地消除扩散的肿瘤细胞,而且更为重要的是,还能提供保护性的预防肿瘤复发的长期免疫记忆功能。02肿瘤疫苗是如何发挥作用的?肿瘤疫苗通过利用肿瘤细胞相关抗原,来唤醒人体针对癌症的免疫系统功能。目标是预防和治疗癌症,或防止癌症的复发。而肿瘤疫苗想在人体中发挥作用,离不开人类MHC(即HLA复合体)的抗原呈递过程。肿瘤疫苗的作用过程为:通过皮下/肌肉注射使抗原进入机体内,被DC细胞摄取。DC细胞通过淋巴管的归巢进入淋巴结,在淋巴结中存在有大量初始T细胞。通过与APC细胞表面的MHC I/II 类分子结合,初始T细胞被激活为CD8或CD4阳性T细胞。激活的T细胞通过血液循环进入肿瘤部位,从而实现对肿瘤细胞的杀伤。T细胞激活与肿瘤细胞杀伤机制[2]03百奥赛图可提供肿瘤疫苗相关临床前安全性及药效评价在进行临床试验之前,疫苗与治疗性药物一样,也需要进行临床前动物安全性评价。临床前动物安全性评价的主要目的是通过相关动物来考察疫苗的安全性,包括对免疫器官和其他毒性靶器官的影响、毒性的可逆性,以及与临床相关的参数,预测其在大规模人群中使用时可能出现的不良反应,降低临床试验受试者和临床使用者承担的风险,并为临床试验方案的制定提供依据。百奥赛图具有完善的临床前药效评价平台,能够从小鼠免疫,肿瘤药效,免疫原性评估提供肿瘤疫苗临床前评价一站式服务,评估抗肿瘤免疫反应。百奥赛图服务能力概览肿瘤药效稳定的肿瘤模型减小对试验设计和疫苗评价的影响,治疗给药与rechallenge实验检测疫苗有效性。 细胞免疫1. 可采用ICS、ELISA、Luminex、MSD等方法检测细胞因子。2. ELISPOT检测T细胞免疫反应Positive Control:加入工作浓度的阳性刺激物;Negative Control:细胞浓度跟实验孔保持一致,不加阳性刺激物;Background:加入不含细胞的培养基,不加阳性刺激物。3. 可用FACS方法检测T/B/NK细胞以及各T细胞亚群比例小鼠肿瘤模型TILS检测与细胞分型生物标记物免疫细胞及小鼠体内标记物4. 此外,我们还能提供体外T细胞杀伤、四聚体检测等服务。体液免疫百奥赛图能够定制化进行肿瘤特异性抗体检测的ELISA方法开发和方法学验证。04百奥赛图上市多款HLA人源化小鼠我们使用HLA人源化小鼠细胞能呈递和识别与人类呈递的表位相似或相同的肽表位。百奥赛图已上市多款HLA人源化小鼠,可助力疫苗领域的研发。下面以B-HLA-A2.1 mice的数据为例:IFN-γ ELISA实验检测B-HLA-A2.1小鼠疫苗诱导的免疫应答。将9-10周龄雌性 B-HLA-A2.1小鼠分为 PBS 组,Group2和Group 3 (n=2)共三组 ,在小鼠双腿内侧肌肉接种 PBS 或疫苗。最后一次免疫三周后处死小鼠。取小鼠脾细胞,用单肽或与靶标无关的多肽作为阴性对照(NC)或anti-mCD3作为阳性对照刺激,然后测定 IFN-γ 的分泌量。各组间体重无显著差异(数据未显示)。(A) 显示用阴性对照、肽疫苗或阳性对照刺激后的免疫小鼠脾细胞。(B) IFN-γ分泌量。结果表明,B-HLA-2.1小鼠为疫苗的体内评价提供了一个强有力的临床前模型。其他相关品系详细信息,可访问百奥动物官网查询或咨询BD。参考资料:[1] Hu Z, Ott PA, Wu CJ. Towards personalized, tumour-specific, therapeutic vaccines for cancer. Nat Rev Immunol. 2018 Mar;18(3):168-182.[2] Hollingsworth RE, Jansen K. Turning the corner on therapeutic cancer vaccines. NPJ Vaccines. 2019 Feb 8;4:7.
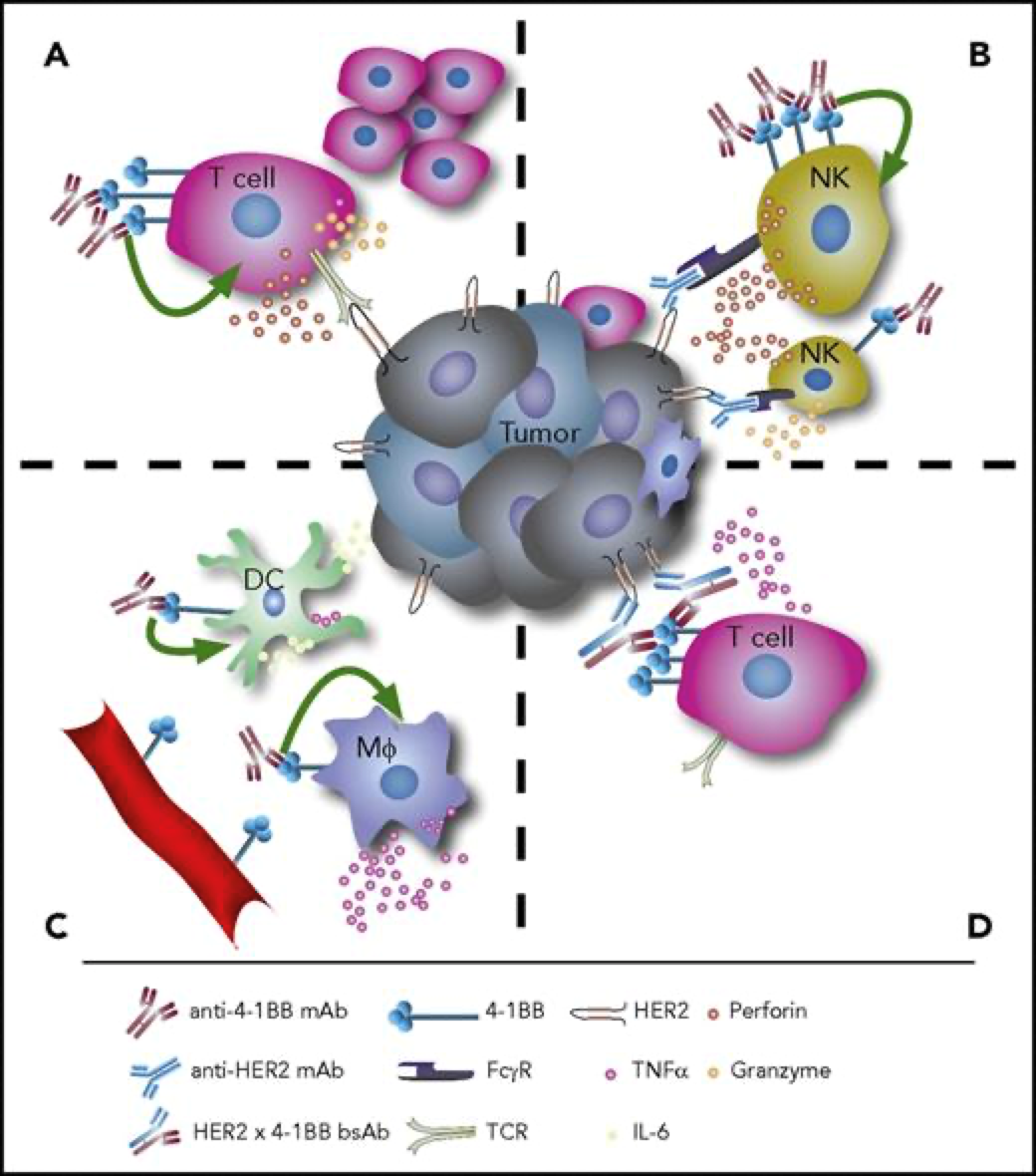
4-1BB靶点介绍4-1BB,即CD137,TNFRSF9(TNF receptor superfamily member 9),它于1989年首次被发现,属于TNF受体超家族。它在抗原启动的T细胞上表达,在静止的T细胞上不表达,除此以外,4-1BB还在树突状细胞(DC)、自然杀伤细胞(NKs)、活化的CD4+和CD8+T淋巴细胞、嗜酸性粒细胞、自然杀伤T细胞(NKT)和肥大细胞中表达。当4-1BB与其配体4-1BBL结合后,可以产生共刺激信号诱导CD4+和CD8+T细胞的活性,促进T细胞增殖,刺激巨噬细胞产生如如IL-6、TNF-α等多种炎症细胞因子。从而激活机体免疫系统,达到抑制肿瘤发生发展、促进肿瘤细胞清除的功效。有研究报道,当在进行4-1BB抗体药物研发时,还要需要考虑抗体结合位点在人和小鼠之间存在的差异性。其结合作用仅限于富含半胱氨酸结构域(CRD),小鼠4-1BBL主要与CRDII结合,人4-1BBL主要与CRDIII结合。小鼠4-1BB/4-1BBL是二聚体,人4-1BB/4-1BBL为三聚体,且人4-1BB不与小鼠的4-1BBL相互作用。[1]图1. 抗4-1BB疗法的免疫调节作用[2]4-1BB药物开发的现状4-1BB作为癌症免疫疗法中十分有潜力的靶点,针对其进行的药物研发非常火热,全球目前已有至少40款4-1BB激动型抗体进入临床研究阶段。4-1BB单抗药物由 BMS研发的Urelumab(全人源IgG4 mAb)是一个4-1BB强激动性的单抗,在I/II期试验中表现出良好的抗肿瘤活性,但在后续治疗过程中发生了两起由肝毒性引起的致命性不良事件。由Pfizer研发的Utomilumab(人源化IgG2 mAb)是一个4-1BB弱激动性的单抗,虽然有较好的肝脏安全性表现,但作为单药治疗抗肿瘤活性十分有限。因此,此前未能很好的在激动作用和肝毒性之间达到良好的平衡,导致4-1BB单抗产品临床应用空间有限;目前新一代单抗设计通过对Fc功能改造试图保留激动性的同时克服安全性问题,Alligator、Agenus以及国内的礼进生物和天演药业的4-1BB单抗产品表现出良好的初步临床数据。4-1BB重点单抗产品临床进度数据来源:Cortellis4-1BB双抗药物平衡抗体激动作用强弱和其肝毒性,是4-1BB单抗药物开发的难点,因此为了克服疗效和毒性的问题,研究者们开始探索新的开发策略。其中以4-1BB为靶点的双抗、三抗以及四抗成为研发的一个热点领域,国内外多家药企均有布局。4-1BB双特异性抗体通过对肿瘤细胞表面靶点和4-1BB亲和力不同精准定位肿瘤微环境,减少肝毒性等副作用。因此双抗疗效相比单抗可以实现更精准的靶向,从而使得临床前的抗肿瘤活性和安全性都得到了提升。如PD-1/PD-L1就是4-1BB双抗常用的另一候选靶点。去年,BioNTech与Genmab联合研发的PD-L1×4-BB双抗药物GEN1046,是一种全长IgG1亚型的PD-L1×4-1BB双抗,其以临床Ⅰ/Ⅱa期结果发表在著名学术期刊《CANCER DISCOVERY》上的文章中,所使用的PD-L1/4-1BB双靶点人源化鼠由百奥动物提供。文中研究结果表明:GEN1046对于CPI耐药的实体瘤治疗效果良好,安全性高,为免疫联合治疗提供了新的方案。4-1BB重点双抗产品临床进度数据来源:Cortellis通过众多临床试验及以往多年研究积累,4-1BB 靶点被证实是肿瘤免疫治疗十分有潜力的靶点,目前如何最大化激动作用并最小化毒性是未来4-1BB激动剂研发亟需解决的问题。百奥动物自主研发的4-1BB系列人源化小鼠(详情见下表),是靶向4-1BB抗体开发相关药物进行药效评价的优质模型。B-h4-1BB/h4-1BBL mice高剂量Urelumab对B-h4-1BB小鼠和B-h4-1BB/h4-1BBL小鼠均产生肝毒性用PBS或Urelumab(内部合成)对纯合子B-h4-1BB小鼠和B-h4-1BB/h4-1BBL小鼠进行治疗 (n=5,雌性,7周龄)。第21天采集血清,检测谷丙转氨酶(ALT)、谷草转氨酶(AST)。与PBS对照相比,使用20mg /kg Urelumab治疗B-h4-1BB小鼠的ALT显著升高,而同样的治疗对B-h4-1BB/h4-1BBL小鼠没有影响。与PBS对照相比,B-h4-1BB小鼠AST无明显升高,但20ug/mL urelumab治疗可使B-h4-1BB/h4-1BBL小鼠AST明显升高。在1mg/kg剂量组,B-h4-1BB小鼠和B-h4-1BB/h4-1BBL小鼠的ALT和AST均无明显变化。表明高剂量urelumab对B-h4-1BB和B-h4-1BB/h4-1BBL小鼠有肝毒性作用。数值以平均值±SEM表示。高剂量Urelumab 治疗可导致B-h4-1BB小鼠和B-h4-1BB/h4-1BBL小鼠肝脏淋巴细胞浸润增加肝毒性的病理分析。在B-h4-1BB小鼠(G1-G3)中,当urelumab剂量为1mg/kg时,肝脏未见明显异常变化(G2)。当剂量为20 mg/kg (G3)时,观察到血管周围细胞浸润或肝脏慢性炎症,并伴有轻度病变。在B-h4-1BB/h4-1BBL小鼠(G4-G6)中,1 mg/kg组(G5) 3/5的小鼠出现病理变化(轻度2/5,轻度1/5),而在20 mg/kg剂量(G6)时,所有实验动物肝脏均出现中度变化。总体而言,20 mg/kg组(G6)肝脏病变程度和发生率显著高于1 mg/kg组(G5)。以上结果提示,高剂量(20mg /kg)的urelumab比低剂量(1mg /kg)的urelumab更容易发生血管周围细胞浸润或肝脏慢性炎症。B-h4-1BB/h4-1BBL小鼠对urelumab毒性作用比B-h4-1BB小鼠更敏感。因此B-h4-1BB/h4-1BBL小鼠是较好的临床前毒性评价模型。B-h4-1BB/h4-1BBL药效验证抗人4-1BB抗体在B-h4-1BB/h4-1BBL小鼠中的抗肿瘤药效。(A) 纯合子B-h4-1BB/h4-1BBL小鼠 (雌性,6-8周龄,n=5) 皮下接种小鼠结肠癌MC38细胞系。结果显示抗人4-1BB抗体能够显著抑制MC38肿瘤细胞的生长。(B)治疗期间体重变化。如图A所示,抗人4-1BB抗体在B-h4-1BB/h4-1BBL小鼠中有效地控制肿瘤生长,这表明B-h4-1BB/h4-1BBL小鼠可为体内评价抗人4-1BB抗体药效提供有力的临床前研究模型。值表示为平均值±SEM。参考文献[1].https://mp.weixin.qq.com/s/kvwfybpx63XQn26xdijIZwhttps://mp.weixin.qq.com/s/999cX0RaXHLyXxZ8MhIaBw[2]. Chester C, Sanmamed MF, Wang J, Melero I. Immunotherapy targeting 4-1BB:mechanistic rationale, clinical results, and future strategies. Blood. 2018 Jan4;131(1):49-57. doi: 10.1182/blood-2017-06-741041. Epub 2017 Nov 8. PMID:29118009.
康洲大数据 Copyright © 2009-2026 药智网YAOZH.COM All Rights Reserved. 工信部备案号:渝ICP备10200070号-3
客户服务热线:400-678-0778 投诉建议:023-62988285转8001 E-mail:admin@yaozh.com
互联网增值电信业务许可证编号:渝B2-20120028 互联网药品信息服务资格证:(渝)-经营性-2021-0017

渝公网安备 50010802001068号

投诉热线: (023) 6262 8397
邮箱: tousu@yaozh.com
QQ: 236960938

