 药智官方微信
药智官方微信

转化生长因子β(TGFβ)是一种多功能的细胞因子,对细胞增殖、分化、粘附、迁移和凋亡具有多重功能。多种类型细胞均可分泌,最早从人血小板中鉴定。哺乳动物中表达三种类型TGFβ:TGFβ1、TGFβ2和TGFβ3,每种由不同的基因编码,并通过结合相同的TGFβ受体发挥相应功能。其中TGFβ2和TGFβ3在胚胎发育中发挥重要作用,而TGFβ1则主要参与控制出生后的免疫反应[1]。三种TGFβ起初均表达为非活性蛋白复合物(SLC或LLC)。TGFβ1同型二聚体与潜伏相关肽(Latent Associated Peptide, LAP)(TGFβ1 N 端具有的长 20~30 个氨基酸的序列)形成的复合被称为小潜复合物 (Small Latent Complex, SLC)。在细胞外,SLC 与 潜伏TGFβ1结合蛋白(Latent TGFβ Binding Protein, LTBP)结合形成的复杂复合物被称为大潜复合物(Large Latent Complex, LLC)。TGFβ的活化有以下3种途径:1)SLC在胞外被蛋白酶水解;2)LLC被细胞外基质铆钉,进而由αβ整合素介导释放有活性的TGFβ;3)SLC被细胞表面的GARP铆钉,并由αβ整合素介导释放有活性的TGFβ。接下来,具有活性的TGFβ与TGFβRII二聚体结合,随后进一步与TGFβRI二聚体结合,形成四聚体,激活下游信号通路,调节基因表达。图1. TGFβ信号通路[2,3]稳态条件下,TGFβ1信号不仅调节多种组织细胞的生长、增殖和分化;也指导免疫系统的耐受和炎症抑制,尤其在胃肠道起重要作用。TGFβ1调节功能的多样性赋予了它在肿瘤发生和发展中亦正亦邪的双面性。在癌症发生早期,TGFβ是一个抑癌因子,可发挥细胞周期阻滞作用,阻断细胞从G1期进入S期,达到抑制肿瘤细胞增殖的作用。然而,随着炎症加剧,一些癌细胞可能会发生突变,使其对TGFβ信号通路不响应,或响应与凋亡脱钩、反变为促进癌细胞转移和定植。同时,肿瘤细胞亦可大量分泌TGFβ1发挥免疫抑制功能,抑制T、B淋巴细胞与NK细胞活化和分化,造成机体免疫功能障碍,使得肿瘤细胞免疫逃逸(图2)。图2. TGFβ的抑癌和促癌信号[2]针对TGFβ信号通路的在研药物主要包括配体抗体、配体陷阱(如bintrafusp alfa)、受配体抗体、小分子激酶抑制剂等,主要聚焦于肿瘤免疫疗法[4]。目前,处于临床阶段的TGFβ1配体抗体药物如下表所示:数据整理自Cortellis数据库为了更好的助力靶向TGFβ1的新药研发,百奥动物自主研发了TGFβ1人源化小鼠B-hTGFB1 mice,用于靶向TGFβ1药物的临床前药效评估。B-hTGFB1 mice 图3. RT-qPCR分析B-hTGFB1小鼠和野生型小鼠的TGFB1 mRNA的表达。B-hTGFB1小鼠中人TGFB1 mRNA表达量与野生型小鼠中的鼠TGFB1 mRNA表达量类似。这表明人TGFB1基因原位替换不影响TGFB1 mRNA的表达。图4. 利用流式细胞仪分析纯合B-hTGFB1小鼠和野生型小鼠的血小板中TGFB1蛋白的种属特异性表达。小鼠的TGFB1仅在野生型小鼠中被检测到,人TGFB1仅在纯合B-hTGFB1小鼠中检测到。脾脏中TGFB1蛋白的表达同样也具有种属特异性(数据未展示)。TGFB1人源化不影响脾脏、淋巴结、血液中白细胞各亚群比例(数据未展示)。图5. 抗体结合实验。从B-hTGFB1小鼠和野生型小鼠取血,用流式细胞术分析血小板人LAP的表达。SRK-181可特异性结合SLC并抑制TGFβ1活化[5]。于是,在SRK-181处理过的纯合B-hTGFB1小鼠和野生型小鼠中均能检测到人LAP的表达;而未处理的鼠中不能检测到人LAP的表达。(hLAP抗体可人鼠交叉识别。)图6. 抗鼠PD-1抗体和抗人TGFβ1抗体联用在B-hTGFB1小鼠中的抗肿瘤作用。(A)mPD-1和hTGFβ1抗体(自制)联用可抑制B-hTGFB1小鼠中MC38肿瘤的生长;(B)治疗期间的体重改变。数值为平均值±SEM。由图可见,抗鼠PD-1抗体和抗人TGFβ1抗体联用可有效控制B-hTGFB1小鼠中的肿瘤生长,说明B-hTGFB1小鼠是TGFβ1抗体药物临床前评估的有力模型。相关产品列表 品系货号B-hTGFB1 mice112245B-hTGFBR2 mice110874B-hGARP mice110102B-hGARP/hTGFB1 mice112241B-hLRRC33 mice110757B-Tgfβ1 cKO mice110164参考资料1. Stockis, J., Dedobbeleer, O. & Lucas, S. Role of GARP in the activation of latent TGF-β1. Molecular bioSystems 13, 1925-1935 (2017).2. Batlle, Eduard, and Joan Massagué. “Transforming Growth Factor-β Signaling in Immunity and Cancer.” Immunity vol. 50,4 (2019): 924-940. doi:10.1016/j.immuni.2019.03.0243. Kelly, Aoife et al. “Regulation of Innate and Adaptive Immunity by TGFβ.” Advances in immunology vol. 134 (2017): 137-233. doi:10.1016/bs.ai.2017.01.0014. Kim, Byung-Gyu et al. “Novel therapies emerging in oncology to target the TGF-β pathway.” Journal of hematology & oncology vol. 14,1 55. 6 Apr. 2021, doi:10.1186/s13045-021-01053-x5. Martin, Constance J et al. “Selective inhibition of TGFβ1 activation overcomes primary resistance to checkpoint blockade therapy by altering tumor immune landscape.” Science translational medicine vol. 12,536 (2020): eaay8456. doi:10.1126/scitranslmed.aay8456

人类白细胞抗原:HLA,通常称之为MHC分子,是在6号染色体短臂上的一类基因。除同卵双生子以外几乎找不到HLA相同者,每个人的HLA千差万别,是识别人类白细胞的重要的分子标志,是免疫系统区分本身和异体物质的基础,具有非常重要的生物学功能。HLA有不同的基因座,编码两大类MHC蛋白。HLA在医学上的意义主要体现在器官移植,输血,疾病相关性,生理学等方面。人6号染色体HLA基因结构HLA复合体有224个基因座(locus),按其产物的结构、分布与功能分为三群。HLA-I经典I类基因: HLA-A、-B、-C参与递呈内源性抗原。非经典I类基因: HLA-E、-G、-F。HLA-II经典II类基因: HLA-DP 、-DQ、-DR参与递呈外源性抗原。非经典II类基因: LMP、TAP、HLA-DM、HLA-DO 参与抗原的加工和转运。HLA-Ⅲ包括编码补体C4、Bf、C2的基因。编码炎症相关分子TNF、HSP70等基因。HLA基因结构HLA基因功能HLA-I可以提供一般细胞内的一些状况,比如该细胞遭受病毒感染,则将病毒外膜蛋白加工成肽链,通过MHC展示到细胞膜表面,供杀手CD8+ T细胞的识别,以进行扑杀。表达于所有有核细胞表面,例外的仅有神经细胞、胰岛外分泌细胞、心肌细胞和精细胞等。HLA-II可以提供细胞外部的情况,像是组织中有细菌侵入,则巨噬细胞进行吞食后,把细菌蛋白加工成肽链,通过MHC展示给辅助性T细胞,启动体液免疫反应。只位于抗原提呈细胞(APC)表面,例如:B细胞、巨噬细胞、树突状细胞、胸腺上皮细胞和人活化的T细胞等。HLA分子对T细胞在胸腺内的分化成熟过程也起重要作用。体外研究发现:去除胸腺中MHCⅡ类抗原阳性的基质细胞,则CD4+ T细胞的发育受阻,在胸腺培养细胞中加入抗MHCⅡ类抗原的单克隆抗体,也能阻止CD4+ T细胞的发育。目前认为MHC分子在T细胞自身耐受的形成和T细胞库的产生中都起着重要作用。HLA-A2.1分子HLA-A2.1是HLA I 类A基因座,第2复等位基因第1号亚型,备受科学家们的关注。关注HLA-A2.1等位基因的原因有很多,下面将介绍其中的一些原因。T细胞的免疫反应依赖于肽与HLA分子的结合,因此研究者经常研究这些分子与各种抗原的相互作用。由于HLA的高度多态性,有成千上万的HLA分子可供选择,为了获得最大的相关性,研究最常见的表达等位基因是最有意义的。HLA-A2.1的肽结合基序早已为人熟知,HLA多聚体可用于抗原特异性T细胞的染色。甚至有小鼠表达HLA-A2.1等位基因,这使得在更接近人类的生物系统中进行临床前测试成为可能。针对HLA-A2.1分子机制研究的需求,BioMice百奥动物自主研发了B-HLA-A2.1 mice和B-NDG HLA-A2.1 mice,助力HLA-A2.1分子研究,为临床前药效评估提供了优质模型。B-HLA-A2.1 mice基本信息蛋白表达分析流式细胞术检测野生型C57BL/6小鼠(+/+)和纯合型B-HLA-A2.1小鼠(H/H)的脾细胞。野生型C57BL/6小鼠可检测到小鼠B2M和H-2Kb/H-2Db。人B2M和HLA-A2.1只在B-HLA-A2.1纯合小鼠中检测到,而小鼠B2M和H-2Kb/H-2Db在B-HLA-A2.1纯合小鼠中检测不到。 野生C57BL/6 (+/+)小鼠和纯合B-HLA-A2.1小鼠 (H/H)注射抗CD3E抗体后,取脾细胞进行流式细胞术检测。结果显示:野生C57BL/6小鼠可检测到小鼠B2M和H-2Kb/H-2Db。纯合B-HLA-A2.1小鼠中只能检测到人B2M和HLA-A2.1,检测不到小鼠B2M和H-2Kb/H-2Db。脾脏白细胞亚群分析从C57BL/6小鼠和纯合B-HLA-A2.1小鼠(n= 3,8周龄,雌性)中分离脾细胞。流式细胞术分析脾细胞以评估白细胞亚群。结果表明:纯合B-HLA-A2.1小鼠的B细胞、树突状细胞、粒细胞、单核细胞和巨噬细胞的百分率与C57BL/6小鼠相似。CD8+ T细胞百分率显著降低,CD4+ T细胞百分率和NK细胞百分率显著升高,说明hB2M-HLA-A2.1-H-2D替代小鼠B2M可能影响CD8+ T细胞的发育,进而影响脾脏T细胞亚型的比例。数值用平均值±SEM表示。在淋巴结和血液中得到同样的结果。脾脏T细胞亚群分析从C57BL/6和纯合B-HLA-A2.1小鼠(n= 3,8周龄,雌性)中分离脾细胞。流式细胞术分析脾细胞以评估T细胞亚群。结果表明:纯合B-HLA-A2.1小鼠中调节性T细胞的百分率与C57BL/6小鼠相似。CD8+ T细胞百分率显著降低,CD4+ T细胞百分率显著升高,说明hB2M-HLA-A2.1-H-2D替代小鼠B2M可能影响CD8+ T细胞的发育,进而影响脾脏T细胞亚型的比例。数值用平均值±SEM表示。在淋巴结和血液中得到同样的结果。B-NDG HLA-A2.1 mice基本信息蛋白表达分析流式细胞术检测B-NDG小鼠(+/+)和纯合B-NDG HLA-A2.1小鼠(H/H)脾细胞。结果表明:小鼠B2M和H-2Kb/H-2Db在B-NDG小鼠中检测到,但在B-NDG HLA-A2.1小鼠中检测不到。人B2M和HLA-A2.1在纯合B-NDG HLA-A2.1小鼠中检测到,而在B-NDG小鼠中未检测到。脾脏白细胞亚群分析从B-NDG小鼠和B-NDG HLA-A2.1小鼠(n= 3,6周龄,雌性)分离脾细胞。流式细胞术分析脾细胞以评估白细胞亚群。结果表明:纯合B-NDG HLA-A2.1小鼠树突状细胞、粒细胞、单核细胞和巨噬细胞的百分率与B-NDG小鼠相似,表明人源化的B2M和HLA-A2.1不改变这些细胞在脾脏的整体发育、分化和分布。数值用平均值±SEM表示。在骨髓和血液中得到同样的结果。人CD34+HSC免疫重建将人CD34+造血干细胞分别植入经0.8 Gy辐照过的B-NDG HLA-A2.1新生小鼠和B-NDG小鼠。(A) B-NDG HLA-A2.1小鼠存活率略低于B-NDG小鼠,但差异无统计学意义。(B)体重。将人CD34+造血干细胞分别植入经0.8 Gy辐照过的B-NDG HLA-A2.1新生小鼠和B-NDG小鼠。流式细胞术检测人白细胞。结果表明,虽然B-NDG HLA-A2.1小鼠的人CD8+ T细胞比例明显低于B-NDG小鼠,但重建24周内人CD8+ T细胞比例仍维持在18%左右。B-NDG HLA-A2.1小鼠人CD4+ T细胞比例明显高于B-NDG小鼠。B-NDG HLA-A2.1小鼠中其他重组细胞类型的比例与B-NDG小鼠相似。参考资料1、Van Laethem, F., Tikhonova, A.N. & Singer, A. MHC restriction is imposed on a diverse T cell receptor repertoire by CD4 and CD8 co-receptors during thymic selection. Trends in immunology 33, 437-441 (2012).2、Garcia, K.C., Adams, J.J., Feng, D. & Ely, L.K. The molecular basis of TCR germline bias for MHC is surprisingly simple. Nature immunology 10, 143-147 (2009).

医药生物行业生命科学领域产业链系列报告显示,随着科研发展与对新靶点的持续追求,基因修饰动物凭借可以精准满足用户需求的特点,正快速发展为模式动物市场主流产品。GMI 数据披露,2019 年全球基因修饰动物模型市场规模约为 100 亿美元,预计 2023 年将增至 141 亿美元,占动物模型服务市场近67%[1]。技术进步重塑市场格局,基因修饰模式动物地位日益凸显。相较于野生型的模式动物,基因修饰模式动物能够对目标基因开展功能缺失或功能获得的研究,达到人类生理或病理更精确的模拟,因此更适合探索研究人类基因功能和人类疾病致病机制,是目前行业重要的发展方向。这其中Cre工具鼠和疾病模型鼠亦扮演着举足轻重的角色。一起围观下百奥动物的蓝宝小家伙带来的好物分享吧~ 鼠界“爱德华”--Cre工具鼠系列INTRODUCTION基因敲除鼠常被用来研究基因的功能,但据研究报道,小鼠有超过30%的基因敲除后导致了胚胎致死或者出生后不久死亡[2],从而无法对该基因在体内发挥的生理功能进行更好的研究。为了解决这个问题,科学家发现了Cre/loxP系统。 1 什么是Cre/loxP系统?Cre-loxp 系统来自于P1 噬菌体,是噬菌体感染病毒实现寄生的“杀手锏”。 Cre重组酶(cyclization recombination enzyme),它是一种由343个氨基酸组成的单体蛋白,可以引发loxP位点的DNA 重组。loxP是一段长34bp的DNA序列,是Cre重组酶识别的位点,被称为loxP位点。Cre重组酶的调控方式有多种多样,有如特异性基因启动子调控的Cre(Promoter-regulated Cre)、配体或药物诱导的Cre(Inducible Cre)、荧光报告的Cre(Fluorescent Cre)以及同时被不同启动子调控的Cre(Split Cre,NCre 和CCre)等。不同调控方式的选择造就了不同的Cre工具鼠。 2 Cre/loxP系统在基因编辑大小鼠中的应用根据Cre-loxp 系统工作的原理,在基因编辑大小鼠中,分别制作Cre重组酶工具大小鼠和带有loxp位点的flox鼠,通过杂交可以实现组织特异性基因敲除,这样可以避免全身性敲除引起的纯合致死情况。你是不是也正在经历实验设计选不到合适Cre鼠的窘境?百奥动物自主开发了一系列Cre工具鼠,助力不同应用场景的科学研究。B-Pgr-iCre小鼠基本信息应用领域生殖系统功能研究表型分析ROSA26-CAG-LSL-tdTomato mice是公司自主研发并验证成功的tdTomato报告小鼠品系。在与Cre小鼠交配后,剔除stop元件,可表达tdTomato红色荧光蛋白。本实验方案利用B-Pgr-iCre mice与CAG-LSL-tdTomato报告小鼠交配,iCre重组酶可介导子代小鼠中表达Pgr的细胞删除LSL元件,表达tdTomato红色荧光蛋白,荧光显微镜下可观察到红色荧光信号,可证明表达Pgr的细胞成功表达iCre重组酶并介导基因重组功能。上图为小鼠不同器官中实验结果的显示,(a)代表小鼠的生殖系统,包括子宫、输卵管和卵巢。(b)代表小鼠的乳腺。IF 检测 tdTomato 红色荧光蛋白和绿色荧光标记 Pgr(progesterone) 的表达。纵轴代表不同基因型的小鼠,横轴代表不同的荧光标记。B-Pgr-iCre(Mut/+);CAG-tdTomato(Mut/+) 小鼠表现出明显的 tdTomato 红色荧光蛋白表达。B-Cdh5-iCreERT2 mice小鼠基本信息应用领域心血管系统,肿瘤和细胞分化机制研究表型分析ROSA26-CAG-LSL-tdTomato mice是公司自主研发并验证成功的tdTomato报告小鼠品系。在与Cre小鼠交配后,剔除stop元件,可表达tdTomato红色荧光蛋白。本次实验中B-Cdh5-iCreERT2 mice与ROSA26-CAG-LSL-tdTomato mice交配,可实现Cdh5-iCreERT2(Mut/+);CAG-LSL-tdTomato(Mut/+)双阳性小鼠在tamoxifen诱导下,在血管淋巴管内皮细胞特异性表达tdTomato红色荧光蛋白。上图为双阳性小鼠主要器官中 tdTomato 和 CD31 的免疫荧光图像。CD31是内皮标记物。(比例尺:25 μm)百奥动物Cre工具鼠产品列表向上滑动查看列表 高能“cosplay”--疾病模型鼠系列INTRODUCTION疾病动物模型是研究人类疾病机制、诊断预后标志物发现、药物筛选和评价等的重要支撑条件。利用基因工程技术对基因进行修饰,可建立敏感动物品系和与人类疾病相同的疾病模型进行药物筛选和药效研究,这类动物可培育为稳定遗传的品系,且具有特定的病理特征,已经成为药物快速筛选的重要手段。百奥动物建立了丰富的疾病模型资源,为疾病治疗药物评价提供稳定有效的模型资源,并能够基于此为广大合作伙伴提供符合国际水平的药理药效服务。血友病模型--B-F8 KO 小鼠F8又称AHF,FVIII(coagulation factor VIII),是位于X染色体上编码凝血因子VIII的基因,在外源性和内源性凝血途径中都不可或缺。它作为凝血因子IXa的辅助因子参与凝血过程,同时借助于Ca2 +和膜磷脂,形成复合物以激活X因子,并进行一系列后续的凝血反应。F8基因的缺失导致严重的凝血障碍,形成伴X染色体隐性遗传的A型血友病。F8基因敲除模型为血友病患者的药物筛选提供了重要的临床前借鉴意义。模型数据验证将实验动物随机分组,分别尾静脉注射给予生理盐水或1mg/kg诺其,给药30min后腹主动脉采血检测血凝指标:活化部分凝血活酶时间 APTT。结果显示:F8 KO小鼠APTT值远高于WT小鼠,给予重组人凝血因子VIIa后,APTT恢复至正常值。结果证明:B-F8 KO小鼠可以作为抗凝血药药效验证的有力工具。高血糖和肥胖症模型--B-ob/ob 小鼠瘦素(Leptin)是由肽链构成的肽类激素。主要由脂肪细胞分泌,其表达主要在白色脂肪组织。此外,在心肌、骨骼肌、胎盘、肺、乳腺上皮和胃黏膜等均有表达。在功能上能够有抑制食欲,增加能量消耗,抑制脂肪合成促进其分解。胰岛素可促进瘦素的分泌,反过来瘦素对胰岛素的合成、分泌发挥负反馈调节。ob是瘦素的编码基因,百奥动物利用基因编辑技术将ob基因2、3号外显子敲除制备的B-ob/ob小鼠,具有肥胖症和高血糖症状,是研究高血糖和肥胖症的有力模型。模型数据验证纯合B-ob/ob小鼠4周龄后体重和血糖(-/-)均持续高于对照组。抗人GCGR抗体药物crotedumab在B-ob/ob小鼠体内的药效。(A)B-ob/ob小鼠的体重变化。(B-C)非空腹血糖和空腹血糖测量。将8-10周龄的雄性B-ob/ob小鼠随机分为2组,每组6-7只。给药组第0天进行给药,第0、3、7天测定非空腹血糖,禁食6小时后测定空腹血糖。(D)Crotedumab可改善葡萄糖耐量。(E)血糖含量曲线下面积。小鼠在自由取水的条件下禁食6h,尾尖取血测定空腹血糖(0 min),腹腔注射葡萄糖2 g/kg,在指定时间测定血糖。(F-G)胰高血糖素和胰岛素测量。结果显示,crotedumab在禁食和非禁食状态下均具有降糖作用,同时能够改善B-ob/ob小鼠的葡萄糖耐量。百奥动物疾病模型鼠产品列表百奥动物可提供多种稳定优质的基因编辑自发疾病动物模型,如肿瘤模型鼠(B-p53 KO 小鼠,B-p53 KO大鼠),高血糖模型鼠(B-ob/ob小鼠),凝血模型鼠(B-F8 KO小鼠,B-F9 KO小鼠)等,提供客户用于相关研究。向上滑动查看产品列表意犹未尽?想要共享蓝宝的更多好物清单,欢迎扫描下方二维码查看或者来电咨询。参考资料:[1]https://mp.weixin.qq.com/s/rhhEFL5A9KhVKAULO29COw[2]Huimin Zhang, Qi Zheng, Ruby Yanru Chen-Tsai;Establishment of a Cre-rat resource for creating conditional and physiological relevant models of human diseases. Transgenic Res. 2021 Feb;30(1):91-104

经历新冠近四年,疫情仍然以星星之火的态势影响着国内外。近两年来,全球研发企业与投资机构的目标几乎都投向了承担着“事前预防”角色的疫苗企业,但此刻亦有许多医药巨头正在悄然行动,在“事后补救”的抗纤维化赛道上积极布局。其中不乏 Boehringer Ingelheim、Genentech、BMS、Roche(Promedior)这些国际大厂,也有 FibroGen、Pliant Therapeutics、Galecto、Blade Therapeutics 等生物技术公司[1]。医药巨头的关注无疑是对领域未来机会的认可。目前,全球仅上市吡非尼酮(抑制TGF-β1的生成和减少血管因子的合成)和尼达尼布(血管生成因子抑制剂)两款药物,但两款药物因其安全性不佳,并且药效有限,仅仅可以延缓病人的纤维化进展,靶点和机理等药理学内容模糊。因此,以 Pfizer 和 Gilead 为代表的跨国制药巨头,已逐渐将新药开发战略重心转移至抗纤维化药物领域中,以满足巨大的市场需求。不是癌症的癌症--纤维化纤维化(Fibrosis)其实是多种类型组织损伤尤其是在慢性炎症性疾病过程中,组织修复反应失调的结局。纤维化几乎发生在所有器官和组织中,例如心、肝、肺、肾、皮肤等,长期以来,人们一直认为纤维化不可逆,但临床前模型和临床试验均表明,它是一个高度动态变化的过程。当组织受到损伤时,多种来源的肌成纤维细胞可通过重塑细胞外环境来启动伤口愈合反应,以恢复组织完整性并促进实质细胞的替换。通常,当组织愈合时,这个促纤维化程序被关闭。然而,持续的损伤和损害会导致这一过程的失调,导致细胞外基质(ECM )蛋白在病理上的过度沉积,并伴随着肌成纤维细胞活性的上调,造成巨噬细胞和免疫细胞浸润的慢性炎症环境。在这种细胞环境中,细胞因子和生长因子被大量释放,包括转化生长因子-β(TGF-β)家族成员和 Wingless/Int-1(Wnt1),它们是纤维化过程的主要效应子。TGF-β 和 Wnt1 结合其干细胞表面受体,并发起下游信号转导,最终分别导致 Smad2/3 和 CBP/β-Catenin 转录调节物的核易位。这导致靶基因表达上调,其功能进一步增强肌成纤维细胞分化和 ECM 蛋白(包括胶原、层粘连蛋白和纤维连接蛋白)的产生和分泌。随着过多 ECM 沉积的进行,基体的结构发生变化并变硬。细胞通过细胞表面整合素受体(激活 Hippo 信号转导通路及其主要下游效应子 YAP 和 TAZ)的机械传导感受 ECM 张力。在另一个前向循环中,激活的 YAP 和 TAZ 转位到胞核,促进包括 CTGF 和 PDGF 在内的促生长基因的上调,这些基因通过 PI3K/AKT/mTOR 通路促进肌成纤维细胞的增殖和活化。纤维化损伤过程[2]常见的与纤维化有关的疾病有:特发性肺纤维化(idiopathic pulmonary fibrosis, IPF)、肝硬化、慢性肾病、心肌梗死、心力衰竭以及非酒精性脂肪肝炎(NASH)等。纤维化还影响肿瘤侵袭和转移、慢性移植物排斥和许多进行性肌病的发病机制。除器官损伤外,纤维化还与癌症进展有关,因为纤维化 ECM 可刺激细胞增殖并改变细胞极性,从而促进肿瘤发育和生长。抗纤维化的应用领域广泛且机制具有一定的共性,因此专注于抗纤维化药企的平台技术具有通用性,目前各家企业的重心一般集中于抗肺纤维化、肝纤维化、肾纤维化、心血管纤维化以及皮肤、视网膜纤维化等领域,其中,领先企业往往先从IPF以及肝纤维化开始,然后进行延展。由于体外药效无法模拟真实体内纤维化环境,抗纤维化药物筛选只能使用漫长的体内试验来进行,几乎所有的新药筛选捷径,包括人工智能,都无法加快抗纤维化药物的研发进程。因此,临床前动物模型在药物研发过程中显得尤为重要,百奥动物自主研发了相应的肝纤维化和肺纤维化模型,为该领域临床前药效评价提供了稳定有效的工具,助力抗纤维化药物研究。肝纤维化模型四氯化碳(CCl4)诱导的肝纤维化四氯化碳(CCl4)诱导的小鼠肝纤维化是被广泛接受的研究肝纤维化实验模型。它在许多方面反映了与毒性损伤相关的人类疾病模式,如α-SMA表达、星状细胞活化和关键基质成分(包括胶原蛋白-1、基质金属蛋白酶及其抑制剂TIMPs)已在该模型的发病机制中得到证实。CCl4诱导肝纤维化模型的建立及组织学评价四氯化碳 (CCl4) 诱导的肝纤维化模型。8周龄雄性 C57BL/6 小鼠腹腔注射浓度为0.25、0.5和0.75 mL/kg的CCl4,每周2次。4、6和8周后小鼠的体重、肝重/体重和血液生化数据如上图。使用CCl4 诱导6周的肝纤维化模型。(A) H&E染色的代表性图片显示 CCl4 诱导后肝脏炎症增加。比例尺:50 μm。(B) 免疫组化的代表性图片,显示肝巨噬细胞(kupffer cells)标记物F480。(C-D) 天狼星红染色的代表性图片,显示肝纤维化增加。比例尺:200 μm。(E-F) 免疫组织化学的代表性图片显示肝脏中的成纤维细胞标志物α-SMA水平。比例尺:300 μm。 奥贝胆酸(OCA)在小鼠肝纤维化模型中的药效验证奥贝胆酸(OCA)治疗后肝纤维化减轻。(A) 图为经CCl4 诱导和 OCA 处理3周后,天狼星红染色显示肝纤维化的代表性图片。(B) 图表示天狼星红染色统计数据。数值为平均值±SEM。*p < 0.05。胆管结扎诱导的肝纤维化胆管结扎引起肝外胆道梗阻,从而导致胆管扩张和胆汁淤积。当胆管内压力进一步升高时,肝内胆管扩张破裂,肝内血管被扩张的胆管和外渗的胆汁同时压迫,肝细胞缺血坏死,纤维组织增生,围绕肝小叶向肝细胞周围扩散,最终可导致肝纤维化。胆管结扎诱导肝纤维化模型的建立胆管结扎肝纤维化模型。(A) 图表示血清中ALT、AST、ALP、GGT和TBIL水平。 (B) 图表示BDL后4周肝脏代表性外观。数值为平均值±SEM。*p<0.05, ***p<0.001, ****p<0.0001。胆管结扎所致肝纤维化模型的组织学评价胆管结扎4周后H&E和天狼星红染色。(A) 图表示H&E染色代表性病理改变。(B) 图天狼星红染色代表性图片显示肝纤维化增加。(C) 图表示胶原纤维阳性信号区比例。数值为平均值±SEM。* p < 0.05。胆管结扎4周后免疫组化染色。 (A-B) 免疫组化染色的代表性图像显示 F4/80 和阳性信号区比例。(C-D) 免疫组化染色代表性图像显示α-SMA水平和阳性信号区比例。硫代乙酰胺(TAA)诱导的肝纤维化模型硫代乙酰胺 (TAA) 是一种广泛用于模拟肝纤维化发病机制中产生损伤的化合物。TAA 通过其代谢产物硫代乙酰胺二氧化硫 (TASO2) 增加活性氧 (ROS) 的形成,引起严重的氧化应激以及脂质过氧化和蛋白质羰基和 DNA 加合物的生成。产生的ROS,引起肝星状细胞 (HSC) 活化,同时诱导造血干细胞转分化为肌纤维母细胞样细胞,导致 EMC 合成和降解失衡及持续纤维化过程。TAA诱导肝纤维化模型的建立及组织学评价TAA 诱导肝纤维化模型4周。(A-B) 血清中 ALT 和 AST 水平。(C) H&E染色的代表性图片。(D-E) 免疫组化染色的代表性图像显示α-SMA水平和阳性信号区比例。(G-F) 天狼星红染色的代表性图像显示肝纤维化增加。TAA 诱导肝纤维化模型6周。(A-B) 血清中 ALT 和 AST 水平。(C) H&E染色的代表性图片。(D-E) 免疫组化染色代表性图像显示α-SMA水平和阳性信号区比例。(F-G) 天狼星红染色的代表性图片,显示肝纤维化增加。肺纤维化模型博来霉素(BLM)诱导的肺纤维化小鼠模型博来霉素 (BLM) 能够引起与接受化疗患者相似的组织学肺模式,是目前诱导动物肺纤维化应用最广泛的药物之一。BLM 通过裂解DNA、诱导炎症反应和增加上皮凋亡剂量发挥其细胞毒性作用,从而刺激肺损伤并导致纤维化。 肺纤维化小鼠模型的建立及组织学评价博来霉素诱导的肺纤维化。C57BL/6J 小鼠在第0天给予博来霉素,每天记录体重,实验结束时收集肺组织,测定肺内羟脯氨酸 (HYP)。 (A) 博来霉素导致体重减轻。 (B)存活率缩短。 (C) HYP 含量增加。 (D) 病理学检查显示显著肺纤维化。 Pamrevlumab 在肺纤维化模型(B-hCCN2小鼠)中的药效验证Pamrevlumab 对博来霉素诱导的体重减轻的影响。B-hCCN2 小鼠在第0天给予博来霉素,然后每两天注射一次 Pamrevlumab 或溶剂,共7次,每天记录体重。实验结果表明,Pamrevlumab治疗能够改善博来霉素诱导的 B-hCCN2 小鼠体重减轻,并可有效降低博来霉素诱导的死亡率。Pamrevlumab 对博来霉素诱导的肺纤维化的影响。 (A) 博来霉素诱导 HYP 含量增加,Pamrevlumab 治疗降低了肺中 HYP 含量。(B) Masson 染色和H&E染色的代表性图像。(C) 条形图显示了纤维化严重程度的定量平均评分。数值为平均值±SEM,n = 4-8,单因素 ANOVA 和 Dunnett 检验,***P < 0.001,**P < 0.01。Micro-CT在肺部疾病模型的研究应用博来霉素诱导肺纤维化模型—直方图、体积根据不同HU区间进行正常、低通气、不通气区域体积的分割。在第4周,结果显示低通气和不通气区域的比例都很高,然后在第10周和第16周之间明显减少,而在正常通气的区域,则呈现相反的趋势[3]。通过使用Micro-CT对小鼠肺部区域进行扫描重建成像,利用肺部分析算法(Hounsfield单位阈值范围分别为-434至-121HU和-120至121HU)实现对通气不良和非通气不良区域的影像区分,从而实现对肺部区域动态检测,并进行功能和结构的评价。量化的影像数据也能提供纵向研究的信息以减少动物之间的变异性,可以更进一步验证体外数据。百奥动物可以提供的模型及检测指标令人期待的未来突破勃林格殷格翰作为肺纤维化治疗领域的全球市场领导者,在这个存在高度未满足医疗需求的领域成功而大胆迈出的第一步是前文所说的尼达尼布(BI BF1120),而今年2月BI 1015550也已经被美国食品药品监督管理局(FDA)授予突破性治疗认定。勃林格殷格翰未来将启动一项III期临床试验,以进一步研究BI 1015550治疗IPF和进行性肺纤维化(PPF)患者的疗效,并希望能尽快让该药物惠及全球患者[4]。Pliant Therapeutics,Inc.是一家临床阶段的生物制药公司,致力于发现和开发用于治疗纤维化的新型疗法。今年7月,Pliant Therapeutics 也公布了其INTEGRIS-IPF IIa期试验积极结果,其药品PLN-74809在患有特发性肺纤维化(IPF)患者身上达到主要与次要终点,呈现剂量相关疗效,并具有良好的耐受性。2021年2月,北京泰德制药与美国Graviton Bioscience Corporation 签订治疗纤维化创新药TDI01海外授权合作协议,首付款及研发、销售里程碑付款最高可达 5.175亿美元。近年来,随着企业临床及各方面的不断推进、纤维化的细胞和分子机制不断被阐明,且新冠疫情后人们对于抗纤维化重视加深,叠加肿瘤赛道的逐渐拥挤,抗纤维化治疗领域被开发的价值不断体现。我们也期待早日拥有更多创新药物用于改善病患生活质量。参考来源:[1] https://mp.weixin.qq.com/s/L8QVMXsh_8-Kk1-iKbeQVA[2] Nie X , Qian L , Sun R , et al. Multi-organ proteomic landscape of COVID-19 autopsies - ScienceDirect[J]. Cell, 2021.[3] Song Shengren,Fu Zhenli,Guan Ruijuan et al. Intracellular hydroxyproline imprinting following resolution of bleomycin-induced pulmonary fibrosis.[J] .Eur Respir J, 2022.[4] https://mp.weixin.qq.com/s/EwbXZN77rPwapjFYdRexVQ

CD20(Cluster of Differentiation 20)是MS4A家族的一员,由297个氨基酸组成的磷蛋白,具有4个跨膜结构域,在B细胞发育中起关键作用。在血液、扁桃体、阑尾、淋巴结和脾脏等多组织表达,正常脑组织和一些免疫细胞上检测不到表达。CD20敲除小鼠与同窝野生型小鼠一样茁壮成长和繁殖,在其出生后第一年未出现任何明显的解剖或形态学异常,或对感染的易感性。CD20没有已知的天然配体,其功能是实现最佳的B细胞免疫应答,特别是针对T细胞非依赖性抗原。CD20表达密度存在B细胞亚群特异性差异,在B细胞淋巴瘤、毛细胞白血病、B细胞慢性淋巴细胞性白血病和黑色素瘤癌干细胞高表达。尽管CD20的功能尚不完全清楚,但它具有动态的细胞活性,是B细胞恶性肿瘤的理想特异性治疗靶标。CD20抗体的6种作用机制[1]CD20抗体CD20抗体是迄今为止最成功的抗肿瘤治疗方案之一,各大药企进行了各种基因工程研究和人源化改造,试图设计和生成强大的CD20抗体。CD20抗体生产的初步成功,推进了单克隆抗体设计方法不断改进。其中4种类型的CD20抗体,包括利妥昔单抗、Ofatumumab、Obinutuzumab和Ublituximab分别于1997年、2010年和2014年在美国和欧洲获批用于治疗慢性淋巴细胞白血病。CD20单克隆抗体设计的不断改进[2]CD20单克隆抗体批准上市时间表及对应症[3]目前,抗 CD20 单抗已经发展到第三代,每一代抗体药物均有各自特有的临床应用价值。01第一代CD20单克隆抗体第一代CD20抗体以利妥昔为代表,针对B细胞上CD20抗原的利妥昔单抗是一种基因工程嵌合鼠/人单克隆抗体,是全球第一款CD20单克隆抗体(mAb)上市药物,上市后迅速成为治疗某些B细胞恶性肿瘤特定类型的标准方案,延续至今。美罗华@利妥昔单抗注射液第一代CD20单抗的疗效很好,甚至对于一直以来的治疗方案产生革命性的影响,但由于其是嵌合鼠/人单克隆抗体,因此使用时会存在一定的排异反应,产生负作用影响治疗效果。为提升治疗效果降低副作用,CD20单抗的药物制备技术不断更新换代,衍生出了第二代CD20单抗。02第二代CD20单克隆抗体第二代抗CD20 mAb将CD20人源化或者全人源改造,降低免疫原性,包括Ofatumumab、Veltuzumab和Ocrelizumab。Ofatumumab (OFA)是一种全人源 I 型抗CD20 IgG1k mAb。Ofatumumab与CD20分子的小和大胞外环(ECL)均结合,在杀伤靶细胞方面比利妥昔单抗更有效。OFA(arzerra)已获批用于治疗氣达拉滨和阿仑单抗治疗失败的复发性或难治性CLL(FA-ref),目前正在开发OFA联合其他药物治疗各种B细胞肿瘤。第二代CD20单克隆抗体[4]与一代CD20抗体相比二代CD20抗体免疫原性降低,减少不良反应,但是二代CD20抗体特异性和抗原结合的亲和力有所下降,因此第三代产品应运而生。03第三代CD20单克隆抗体第三代人源化CD 20 mAb具有Fc工程化改造,以增加其与FcyRIIIa受体的结合亲和力,包括 Ocaratuzumab、PR0131921和Obinutuzumab,均在进行不同适应症的临床开发。Ocaratuzumab(AME-133v)是一种 I 型人源化IgG1 mAb。其与CD20的结合亲和力增加13-20倍,与FcyRIIIa受体低亲和力(F/F和F/V)变体的亲和力增加5-7倍。这些可能是克服利妥昔单抗缓解率较低和缓解持续时间较短的机制。Obinutuzumab(GA101)在2013 年已获得美国 FDA 批准上市,主要适用于未经治疗的慢性淋巴细胞白血病患者以及联合苯达莫司汀二线治疗复发性滤泡性淋巴瘤。第三代CD20单抗[4]虽然CD20单抗的药物制备技术经过多次更新换代,二代三代各具优势,但与一代产品利妥昔单抗相比,在适应症和疗效上并未更具优势,因此目前市场上第一代产品利妥昔单抗依然占据绝对地位。发展前景CD20 单抗是世界上第一个上市的单克隆抗体,具有多年的临床治疗数据,对应不同的适应症以及设计优化,该靶点尚具有巨大的市场潜力。此外基于CD20靶点的研究的人源化单克隆抗体、双特异性抗体、CAR-T疗法、抗体偶联药物(ADC)、与小分子药物联合治疗等研究,均取得良好的效果。国内外有多家企业展开针对CD20靶点的单抗,双抗以及联用药物的研究,超110多个研究管线处于临床前期或已进入临床阶段,超23款药物已上市。今年9月,北京神州细胞自主研发的新型抗CD20单抗——“安平希”瑞帕妥单抗获批上市,是国产首款抗CD20单抗上市药物。国内外部分CD20药物临床研发进展数据来源:Cortellis通过众多临床试验及以往多年研究积累,证实了 CD20 靶点是治疗 B 细胞淋巴瘤的重要靶点,以CD20 为靶点研发出的药物在临床治疗 B 细胞淋巴瘤中发挥了关键作用。百奥动物自主研发的CD20系列人源化小鼠及细胞系,是靶向CD20抗体开发相关药物进行药效评价的优质模型。B-hCD20 mice01基本信息02mRNA表达分析RT-PCR分析CD20基因在野生型小鼠和B-hCD20小鼠中的表达情况。鼠CD20 mRNA仅在野生型(+/+)小鼠脾细胞中可检测到。人CD20 mRNA仅在纯合子B-hCD20(H/H)小鼠中检测到,而在野生型(+/+)小鼠中检测不到。03蛋白表达分析 用流式细胞术分析B-hCD20纯合子小鼠CD20的蛋白表达。采集野生型C57BL/6小鼠和纯合子B-hCD20(H/H)小鼠脾细胞,用种属特异性抗CD20抗体进行流式细胞术分析。结果显示:鼠CD20仅在野生型C57BL/6(+/+)小鼠中检测到;人CD20仅在纯合子B-hCD20(H/H)小鼠中检测到,在C57BL/6(+/+)小鼠中检测不到。B-hCD28/hCD20 mice01基本信息02蛋白表达分析流式细胞术分析B-hCD28/hCD20纯合子小鼠CD28的蛋白表达。采集野生型C57BL/6(+/+)小鼠和纯合子B-hCD28/hCD20(H/H;H/H)小鼠脾细胞,用种属特异性抗CD28抗体进行流式细胞术分析。结果显示:鼠CD28仅在C57BL/6(+/+)小鼠中检测到;人CD28仅在纯合子B-hCD28/hCD20(H/H;H/H)小鼠中检测到,在C57BL/6(+/+)小鼠中检测不到。流式细胞术分析B-hCD28/hCD20纯合子小鼠CD20特异性表达。采集野生型C57BL/6(+/+)小鼠和纯合B-hCD28/hCD20 (H/H;H/H)小鼠脾细胞,用种属特异性抗CD20抗体进行流式细胞术分析。结果显示:鼠CD20仅在C57BL/6(+/+)小鼠中检测到;人CD20仅在纯合子B-hCD28/hCD20(H/H;H/H)小鼠中检测到,在C57BL/6(+/+)小鼠中检测不到。B-hCD3E/hCD20 mice01基本信息02蛋白表达分析流式细胞术分析B-hCD3E/hCD20纯合子小鼠CD3E的蛋白表达。采集野生型C57BL/6(+/+)小鼠和纯合子B-hCD3E/hCD20(H/H;H/H)小鼠脾细胞,用种属特异性抗CD3E抗体进行流式细胞术分析。结果显示:鼠CD3E仅在C57BL/6(+/+)小鼠中检测到;人CD3E仅在纯合子B-hCD3E/hCD20(H/H;H/H)小鼠中检测到,在C57BL/6(+/+)小鼠中检测不到。流式细胞术分析B-hCD3E/hCD20纯合子小鼠CD20的蛋白表达。采集野生型C57BL/6(+/+)小鼠和纯合子B-hCD3E/hCD20(H/H;H/H)小鼠脾细胞,用种属特异性抗CD20抗体进行流式细胞术分析。结果显示:鼠CD20仅在C57BL/6(+/+)小鼠中检测到;人CD20仅在纯合子B-hCD3E/hCD20(H/H;H/H)小鼠中检测到,而在C57BL/6(+/+)小鼠中检测不到。03免疫分型脾脏白细胞亚群的流式分析。分离野生C57BL/6(+/+)小鼠和B-hCD3E/hCD20 (H/H;H/H)小鼠(雌性,n=3,6周龄)的脾细胞并进行白细胞亚群流式分析。A.代表性单活CD45+细胞的流式分析。B.图A的统计学分析。结果显示:纯合子B-hCD3E/hCD20小鼠的T细胞、B细胞、NK细胞、树突状细胞、粒细胞、单核细胞和巨噬细胞的比例与C57BL/6小鼠相似,证明人源化CD3E/CD20不影响脾脏中这些细胞集群总体的发育、分化或分布。数值:平均值 ± SEM脾脏T细胞亚群的流式分析。分离野生型C57BL/6(+/+)小鼠和B-hCD3E/hCD20 (H/H;H/H)小鼠(雌性,n=3,6周龄)的脾细胞并进行T细胞亚群流式分析。A.代表性CD3+ T细胞的流式图分析。B. 图A的统计学分析。结果显示:纯合子B-hCD3E/hCD20小鼠的CD8+ T细胞、CD4+ T细胞和Treg细胞的比例与C57BL/6小鼠相似,证明人源化CD3E/CD20不影响脾脏中这些T细胞集群总体的发育、分化和分布。数值:平均值 ± SEM人源化CD3E/CD20同样也不影响淋巴结、血液中白细胞亚群和T细胞亚群总体的发育、分化和分布。(数据未展示)B-hCD3EDG/hCD20 mice01基本信息02mRNA表达分析RT-PCR分析CD3DG基因在野生型小鼠和B-hCD3EDG/hCD20小鼠中的表达。小鼠Cd3d和Cd3g mRNA仅在野生型(+/+)小鼠胸腺细胞中检测到。人CD3D和CD3G mRNA仅在纯合B-hCD3EDG/hCD20(H/H;H/H)小鼠中检测到,而在野生型(+/+)小鼠中检测不到。03蛋白表达分析流式细胞术分析纯合B-hCD3EDG/hCD20小鼠中CD3E的蛋白表达。采集野生型C57BL/6 (+/+)小鼠和纯合子B-hCD3EDG/hCD20(H/H;H/H)小鼠脾细胞,用种属特异性抗CD3E抗体进行流式细胞术分析。结果显示:鼠CD3E仅在C57BL/6 (+/+)小鼠中检测到;人CD3E仅在纯合子B-hCD3EDG/hCD20 (H/H;H/H)小鼠中检测到,而在C57BL/6 (+/+)小鼠中检测不到。流式细胞术分析B-hCD3EDG/hCD20纯合子小鼠中CD20的蛋白表达。采集野生型C57BL/6 (+/+)小鼠和纯合子B-hCD3EDG/hCD20 (H/H;H/H)小鼠脾细胞,采用种属特异性抗CD20抗体进行流式细胞术分析。结果显示:鼠CD20仅在C57BL/6 (+/+)小鼠中可检测到;人CD20仅在纯合子B-hCD3EDG/hCD20(H/H;H/H)小鼠中检测到,而在C57BL/6 (+/+)小鼠中检测不到。CD20靶点相关模型列表参考文献:1. Payandeh, Z. et al. The applications of anti-CD20 antibodies to treat various B cells disorders. Biomed Pharmacother 109, 2415-2426 (2019).2. Lim, S.H. et al. Anti-CD20 monoclonal antibodies: historical and future perspectives. Haematologica 95, 135-143 (2010).3. Marshall, M.J.E., Stopforth, R.J. & Cragg, M.S. Therapeutic Antibodies: What Have We Learnt from Targeting CD20 and Where Are We Going? Front Immunol 8, 1245 (2017).4. Shundong Cangl, N.M., Kemeng Wang2 and Delong Liu2. Novel CD20 monoclonal antibodies for lymphoma therapy. Cang et al. Journal of Hematology & Oncology 5 (2012).

近年来,免疫治疗在抗肿瘤治疗中的作用备受关注,基于免疫治疗的新药开发和标志物探索,成为了当前肿瘤研究的热点,这也对临床前研究动物模型的建立提出了更高的要求。即可以模拟人肿瘤特征又同时存在“人源化”免疫系统的免疫重建小鼠模型,成为了免疫肿瘤研发中的优质模型。百奥动物B-NDG小鼠缺乏成熟的T、B、NK细胞,是目前国际公认的免疫缺陷程度高、适合人源细胞或组织移植的工具小鼠。将人的免疫细胞、造血干细胞移植到B-NDG及B-NDG衍生小鼠中构建的免疫系统重建小鼠,能够更好的模拟人的免疫系统,进行免疫学研究和免疫药物评价。但在使用重度免疫缺陷小鼠进行PBMC和CD34+ HSC免疫重建时会面临一些常见问题,如PBMC重建后的T细胞会对小鼠自身细胞进行攻击导致严重的xeno-GvHD反应;CD34+ HSC重建后的NK细胞和髓系细胞重建比例不足。我们以B-NDG小鼠为基础分别开发了可延缓xeno-GvHD反应和可促进髓系细胞发育的二代系列小鼠,以满足特定细胞功能研究和相应靶点药物评价的需求。减轻GvHD反应,延长实验窗口期B-NDG MHC I/II DKO mice plus 不表达MHC I/II 类分子B-NDG小鼠、B-NDG B2m KO plus小鼠和B-NDG MHC I/II DKO plus小鼠进行人PBMC重建后GvHD严重程度的比较 第0天将3个健康供体的人PBMCs (5E6)静脉注射B-NDG小鼠、B-NDG B2m KO plus小鼠和B-NDG MHC I/II DKO plus小鼠(雌性,5周龄,n=5)中。A.用Kaplan Meier生存曲线分析小鼠存活率。B.体重变化。GvHD临床体征每周评分两次。结果表明:B-NDG MHC I/II DKO plus小鼠可显著延缓GvHD发生,并降低GvHD严重程度。因此B-NDG MHC I/II DKO plus小鼠更适合将人PBMC移植到免疫缺陷小鼠模型。B-NDG B2m KO mice plus B-NDG小鼠、B-NDG B2m KO plus进行人PBMC重建后GvHD严重程度的比较将3个健康供体的2E6人PBMC静脉注射B-NDG B2m KO plus小鼠(雌性,11周龄,n=5)和B-NDG小鼠(雌性,10周龄,n=6)中。GvHD临床体征每周评分两次。结果表明:在人PBMC诱导的GvHD模型中,B-NDG B2m KO plus小鼠比B-NDG小鼠生存期明显延长,B-NDG B2m KO plus小鼠可延缓GvHD的发病并减轻其严重程度。促进髓系细胞重建B-NDG MGMT3 mice 人源化IL3, CSF2, CSF1, THPO 基因未经辐照的B-NDG MGMT3小鼠进行人CD34+HSC重建将人CD34+HSC造血干细胞(3E4)静脉注射B-NDG小鼠和B-NDG MGMT3小鼠(出生后24-72h,n=15)。B-NDG小鼠给予1.0 gy辐照,B-NDG MGMT3小鼠不辐照。流式细胞分析两种小鼠进行人CD34+HSC免疫重建后的外周血淋巴细胞。结果表明:未经辐照的B-NDG MGMT3小鼠的CD45+细胞比例从移植后12周开始达到25%,并持续上升,明显高于B-NDG小鼠。B-NDG MGMT3小鼠单核细胞、MDSCs、DCs和Treg的比例高于B-NDG小鼠。(注:A面板第18周数据因流式细胞检测问题无意义。)B-NDG hCSF1/hTHPO mice 未经辐照的B-NDG hCSF1/hTHPO小鼠进行人CD34+HSC重建将人CD34+HSC细胞(0.15 M)静脉注射到纯合B-NDG hCSF1/hTHPO小鼠(雌性,6周龄,n=15)。流式细胞术分析人CD34+HSC免疫重建后小鼠外周血淋巴细胞。结果显示:未经辐照的B-NDG hCSF1/hTHPO小鼠成功重建了T、包括B、NK、髓系细胞、单核细胞和中性粒细胞在内的人多系细胞。免疫缺陷动物产品列表

多发性骨髓瘤(Multiple Myeloma, MM)是全球第二大常见的恶性血液肿瘤,始于骨髓中健康的浆细胞恶性增殖。在当前是一种较难治愈的疾病,大多数患者终将复发。据统计,2020年全球有超过17万人被诊断出患有多发性骨髓瘤,60岁以上老人属于该病的高发群体,发病年龄亦有呈年轻化的趋势。随着我国人口老龄化趋势上升,多发性骨髓瘤的防治负担将逐年加剧,成为危害人们身体健康的一大挑战。多发性骨髓瘤从癌前到有明显疾病症状的演变过程,在SMM阶段的早期发现和早期干预以及预防或治疗策略可能是提高这种复杂疾病治愈率的途径。[1] 左图:正常的骨髓 ,右图:多发性骨髓瘤;多发性骨髓瘤中可见大量恶性浆细胞,其特征是在细胞核附近的细胞质内有一个苍白的区域。[2]目前,针对MM的治疗药物有糖皮质激素、细胞毒性药物、免疫抑制剂、蛋白酶抑制剂、单抗和细胞疗法等。其中,免疫治疗已经在许多癌症领域被证明是革命性疗法,但由于MM存在免疫抑制微环境现象,影响了免疫治疗的疗效,导致其对MM的治疗进展较为缓慢。MM细胞与骨髓(BM)微环境之间的相互作用,对MM的发病起着关键作用,可通过诱导或分泌细胞因子促进肿瘤细胞生长、免疫逃逸及耐药,增加Treg细胞的数量,抑制效应T细胞的杀伤等。靶向特定肿瘤抗原或逆转具有免疫抑制作用的骨髓微环境的免疫疗法可以帮助改善MM的标准治疗方案。骨髓MM微环境中的抗骨髓瘤作用[3]细胞重定向BiAb和BiTE同时结合MM细胞上的骨髓瘤特异性抗原和T细胞上的CD3。MM抗原包括BCMA, CD38, CS1/SLAMF7, GPRC5D和FcRH5,如图示。BiAbs或自然杀伤细胞衔接器(NKCEs)也靶向自然杀伤(NK)细胞相关受体抗原(如CD16A, NKG2D, NKp30),激活NK细胞并增强其抗MM活性。双特异性分子,包括双特异性抗体(BsAbs)和双特异性T细胞衔接器(BiTEs),通过同时结合MM细胞和免疫效应细胞上的抗原,使这些细胞靠近从而促进免疫细胞裂解MM细胞。靶向BCMA和GPRC5D的BsAbs已展示出很好的临床疗效,针对FcRH5的早期临床试验结果也非常有前景。这些药物的免疫调节作用不依赖主要组织相容性复合体(MHC) I类的抗原提呈,可在没有共刺激的情况下发生,适用于免疫系统功能失调的MM患者。BCMA01全称B细胞成熟抗原,又名CD269或TNFRSF17,属于肿瘤坏死因子受体超家族成员,仅高表达于浆细胞表面,部分表达于浆细胞样树突状细胞,是MM免疫治疗的理想靶点。针对BCMA靶点开发的肿瘤免疫疗法主要分为3类:嵌合抗原受体T细胞疗法(CAR-T)、双特异性抗体(BsAb)、抗体药物偶联物(ADC)。GPRC5D02全称G蛋白偶联受体C57家族亚型D,为7次跨膜蛋白,高表达于浆细胞表面,低表达于毛囊区域,其他健康细胞则不表达,为治疗MM的潜在候选靶点。GPRC5D的表达与BCMA不相关,联合靶向这两个靶点的疗法可以发挥互补效应或开发双靶点CAR-T、双抗。FcRH503又称FcRL5、CD307或免疫球蛋白超家族受体易位相关蛋白2 (Immunoglobulin Superfamily Receptor Translocation Associated 2, IRTA2),是一种功能未知的膜蛋白,只表达于B细胞系,包括骨髓瘤细胞。临床前研究的数据表明,FcRH5/CD3双抗可成功激活T细胞,诱导细胞因子产生,并清除恶性浆细胞。根据科睿唯安数据库检索,靶向MM相关靶点药物研究数量众多,部分靶点研究进展情况见下表。强生的Darzalex是在2015年第一批被批准用于MM治疗的免疫疗法,这一时期MM的抗体药研发主要集中在靶向CD38靶点;到2020年GSK研发的首个靶向BCMA药物Blenrep在美获批,2021年BMS的Abecma上市,今年传奇生物的Carvykti上市以及强生的Teclistamab作为全球首个CD3/BCMA双抗获批即将上市,另外还有众多管线处在临床前或临床试验阶段,靶向BCMA赛道的药物研发可谓相当火热。布局靶向MM治疗药物的新靶点,避免同质化竞争,或许也不失为后来者开发该适应症药物以占得先机的上策,当然面临的风险也随之提高。部分药物研究进展整理自科睿唯安数据库及网络百奥动物利用基因编辑技术自主开发了MM相关靶点人源化小鼠及细胞系,助力抗体药物临床前研究。部分数据展示如下:B-hCD38 mice蛋白表达分析采集野生型C57BL/6小鼠和纯合B-hCD38小鼠的脾细胞和血液,用种特异性抗CD38抗体进行流式细胞术分析。小鼠CD38在WT小鼠中检测到。纯合B-hCD38中只检测到人CD38,而WT小鼠检测不到人CD38。药效验证将小鼠T淋巴细胞瘤B-hCD38-luc E.G7-OVA细胞经尾静脉注射到B-hCD38纯合小鼠(雌性,6周龄,n=6)体内。当总通量达到约106 Ig时,将小鼠分组,并用抗人CD38抗体对其进行治疗。(A) 抗人CD38抗体(内部合成)抑制B-hCD38-luc E.G7-OVA小鼠肿瘤生长。(B) 治疗期间体重变化。(C) B-hCD38-luc E.G7-OVA细胞的体内荧光素酶成像图。每周2次测量信号强度和体重,第0、3、7、10天行影像学检查。值表示为平均值±SEM。B-hBCMA micemRNA表达分析RT-PCR分析野生型C57BL/6小鼠和B-hBCMA小鼠BCMA基因特异性表达情况,鼠Bcma mRNA仅在野生型C57BL/6小鼠脾细胞中检测到,人BCMA mRNA仅在纯合B-hBCMA小鼠中检测到。B-hGPRC5D micemRNA和蛋白表达分析(A) RT-PCR分析野生型C57BL/6小鼠和B-hGPRC5D小鼠GPRC5D基因的特异性表达情况,鼠Gprc5d mRNA仅在野生型C57BL/6小鼠睾丸中检测到。人GPRC5D mRNA仅在纯合B-hGPRC5D小鼠中检测到。(B) 取野生型C57BL/6小鼠和纯合B-hGPRC5D小鼠的脾脏,用抗GPRC5D抗体进行western blot分析。由于抗体的交叉反应性,GPRC5D在WT小鼠和纯合B-hGPRC5D小鼠中均可检测到。 相关产品列表 更多数据信息,欢迎联系我们。参考资料[1] https://doi.org/10.1016/j.ctrv.2021.102284[2] https://www.cancer.net/cancer-types/multiple-myeloma/introduction[3] doi: 10.3389/fonc.2022.1032775[4] doi:10.3390/jcm9072166
嗜酸性胃炎(EG)、嗜酸性十二指肠炎(EoD)和嗜酸性食管炎是一类严重的炎症性疾病,表现为胃、十二指肠和食管外嗜酸性粒细胞水平升高。目前没有针对此种疾病的有效疗法,采用全身性的类固醇激素疗法能够改善疾病症状,但长期使用会带来严重副作用。SIGLEC8因为具有强大的嗜酸性粒细胞促凋亡和肥大细胞抑制活性,并选择性表达在这些关键的炎症效应细胞的作用,被认为是很有前途的抗炎靶点。靶点概述Siglecs(结合唾液酸的免疫球蛋白型凝集素)是主要在造血系统细胞上发现的一类免疫调节受体。迄今为止,已在人类中鉴定出14种SIGLEC,主要在各种白细胞群上表达。[1]根据进化上的保守程度分为进化保守型和非保守型两类SIGLEC。保守型SIGLEC由SIGLEC1、SIGLEC2、SIGLEC4和SIGLEC15组成,它们在哺乳动物中存在直系同源基因;非保守型SIGLEC则由其他成员组成,在哺乳动物物种中并不存在真正的同源基因,被称为CD33相关基因 SIGLEC,其中就包括SIGIEC8。图1. SIGLEC8促进嗜酸性粒细胞凋亡并减少肥大细胞炎症反应&SIGLEC8的潜在聚糖配体[2]SIGLEC8是属于SIGLEC 家族 CD33样亚家族成员,全称为“sialic acid binding Ig like lectin 8”。它仅在人类嗜酸性粒细胞和肥大细胞上高度表达,在嗜碱性粒细胞上表达较弱。在体外,SIGLEC8在嗜酸性粒细胞上与抗体或合成聚糖配体包被的聚合物交联可诱导其快速凋亡,而在肥大细胞上,SIGLEC8 连接可抑制 IgE/FceRI介导的炎症介质释放,而不影响他们的生存[3]。用促进存活的促炎性细胞因子预激活嗜酸性粒细胞(过敏性炎症的一个关键过程)不仅不能抵消 SIGLEC8 触发的细胞死亡,反而会有效增强其对 SIGLEC8连接反应中细胞凋亡的敏感性,这表明SIGLEC8介导的免疫抑制在炎症条件下发挥着特殊作用,被认为是很有前途的抗炎靶点,因此在治疗过敏反应、哮喘等由这些细胞引发的炎症性疾病方面具有重要作用[4]。新药研究进展目前以SIGLEC8为靶点的在研药物共5项,开发机构均是Allakos。其中Antolimab(AK-002)处于临床II期,其余四项处于临床前阶段。Antolimab,是一款靶向SIGLEC8的人源重组非岩藻糖基化单克隆IgG1抗体的药物;能够通过SIGLEC8移植肥大细胞和消耗嗜酸性粒细胞,从而抑制有这两类细胞的错误激活导致的一系列疾病,如嗜酸性胃炎(EG)、嗜酸性十二指肠炎(EoD)和嗜酸性食管炎等慢性炎症疾病。2020年5月Allakos公布Antolimab在治疗EG、EoD和嗜酸性食管炎上已经获得了 FDA的孤儿药认定资格。数据来源于科睿唯安为助力靶向SIGLEC8的新药研发,百奥赛图成功开发了相应靶点人源化小鼠。B-hSIGLEC8 miceB-hSIGLEC8 小鼠中的嗜酸性粒细胞和肥大细胞与抗人 SIGLEC8 抗体结合通过流式细胞术分析 B-hSIGLEC8 小鼠的嗜酸性粒细胞和肥大细胞。从野生型 C57BL/6 小鼠和B-hSIGLEC8小鼠中分离出嗜酸性粒细胞和腹膜灌洗肥大细胞。使用基准抗体 Lirentelimab(内部合成)进行流式细胞术分析以评估人 SIGLEC8 表达。Lirentelimab 在纯合 B-hSIGLEC8 小鼠中仅与 SIGLEC8结合,在野生型小鼠中不与 SIGLEC8结合。小鼠哮喘模型BALF免疫细胞数量从B-hSIGLEC8小鼠(n=6)中分离出BALF免疫细胞。在PBS或lirentelimab(内部合成)处理下,通过流式细胞术分析嗜酸性粒细胞的数量。lirentelimab 治疗后,纯合 B-hSIGLEC8 小鼠中炎症细胞的表达水平低于阳性对照。抗人SIGLEC8抗体的药效B-hSIGLEC8 小鼠哮喘样模型中的 H&E 染色。(A) 在研究结束时收集肺组织。H&E染色结果显示,暴露于PBS气雾剂的B-hSIGLEC8小鼠的肺组织没有表现出任何炎症。暴露于OVA导致 B-hSIGLEC8 小鼠的支气管周围和血管周围炎症显着增加。(B) 在用 lirentelimab(内部合成)治疗的小鼠中观察到嗜酸性粒细胞浸润显着减少。注:治疗模型组中lirentelimab单抗通过气管内雾化给药。本期分享到此结束,如果您对相关小鼠模型有需求,欢迎随时与我们联系。参考文献[1] Ravetch JV, Lanier LL (2000) Immune inhibitory receptors.Science290(5489):84–89.[2] Angata T, Varki A. Discovery, classification, evolution and diversity of Siglecs. Mol Aspects Med. 2023 Apr;90:101117. doi: 10.1016/j.mam.2022.101117.Epub 2022 Aug 18. PMID: 35989204; PMCID: PMC9905256.[3] Hudson SA, Bovin NV, Schnaar RL, Crocker PR, Bochner BS (2009) Eosinophil-selectivebinding and proapoptotic effect in vitro of a synthetic Siglec-8 ligand, polymeric6′-sulfated sialyl Lewis x.J Pharmacol Exp Ther330(2):608–612.[4] Na HJ, Hudson SA, Bochner BS (2012) IL-33 enhances Siglec-8 mediated apoptosis ofhuman eosinophils.Cytokine57(1):169–174.
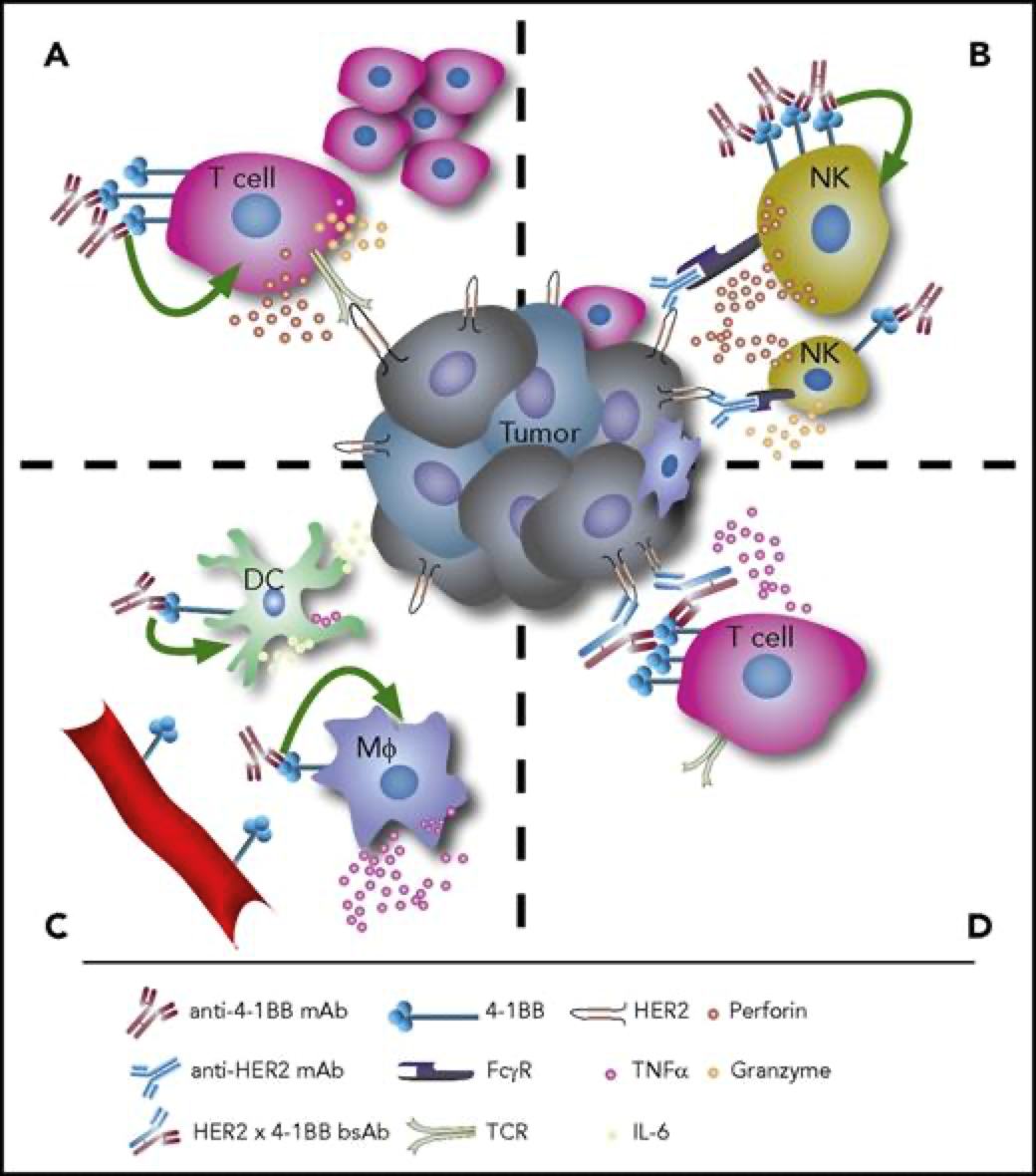
4-1BB靶点介绍4-1BB,即CD137,TNFRSF9(TNF receptor superfamily member 9),它于1989年首次被发现,属于TNF受体超家族。它在抗原启动的T细胞上表达,在静止的T细胞上不表达,除此以外,4-1BB还在树突状细胞(DC)、自然杀伤细胞(NKs)、活化的CD4+和CD8+T淋巴细胞、嗜酸性粒细胞、自然杀伤T细胞(NKT)和肥大细胞中表达。当4-1BB与其配体4-1BBL结合后,可以产生共刺激信号诱导CD4+和CD8+T细胞的活性,促进T细胞增殖,刺激巨噬细胞产生如如IL-6、TNF-α等多种炎症细胞因子。从而激活机体免疫系统,达到抑制肿瘤发生发展、促进肿瘤细胞清除的功效。有研究报道,当在进行4-1BB抗体药物研发时,还要需要考虑抗体结合位点在人和小鼠之间存在的差异性。其结合作用仅限于富含半胱氨酸结构域(CRD),小鼠4-1BBL主要与CRDII结合,人4-1BBL主要与CRDIII结合。小鼠4-1BB/4-1BBL是二聚体,人4-1BB/4-1BBL为三聚体,且人4-1BB不与小鼠的4-1BBL相互作用。[1]图1. 抗4-1BB疗法的免疫调节作用[2]4-1BB药物开发的现状4-1BB作为癌症免疫疗法中十分有潜力的靶点,针对其进行的药物研发非常火热,全球目前已有至少40款4-1BB激动型抗体进入临床研究阶段。4-1BB单抗药物由 BMS研发的Urelumab(全人源IgG4 mAb)是一个4-1BB强激动性的单抗,在I/II期试验中表现出良好的抗肿瘤活性,但在后续治疗过程中发生了两起由肝毒性引起的致命性不良事件。由Pfizer研发的Utomilumab(人源化IgG2 mAb)是一个4-1BB弱激动性的单抗,虽然有较好的肝脏安全性表现,但作为单药治疗抗肿瘤活性十分有限。因此,此前未能很好的在激动作用和肝毒性之间达到良好的平衡,导致4-1BB单抗产品临床应用空间有限;目前新一代单抗设计通过对Fc功能改造试图保留激动性的同时克服安全性问题,Alligator、Agenus以及国内的礼进生物和天演药业的4-1BB单抗产品表现出良好的初步临床数据。4-1BB重点单抗产品临床进度数据来源:Cortellis4-1BB双抗药物平衡抗体激动作用强弱和其肝毒性,是4-1BB单抗药物开发的难点,因此为了克服疗效和毒性的问题,研究者们开始探索新的开发策略。其中以4-1BB为靶点的双抗、三抗以及四抗成为研发的一个热点领域,国内外多家药企均有布局。4-1BB双特异性抗体通过对肿瘤细胞表面靶点和4-1BB亲和力不同精准定位肿瘤微环境,减少肝毒性等副作用。因此双抗疗效相比单抗可以实现更精准的靶向,从而使得临床前的抗肿瘤活性和安全性都得到了提升。如PD-1/PD-L1就是4-1BB双抗常用的另一候选靶点。去年,BioNTech与Genmab联合研发的PD-L1×4-BB双抗药物GEN1046,是一种全长IgG1亚型的PD-L1×4-1BB双抗,其以临床Ⅰ/Ⅱa期结果发表在著名学术期刊《CANCER DISCOVERY》上的文章中,所使用的PD-L1/4-1BB双靶点人源化鼠由百奥动物提供。文中研究结果表明:GEN1046对于CPI耐药的实体瘤治疗效果良好,安全性高,为免疫联合治疗提供了新的方案。4-1BB重点双抗产品临床进度数据来源:Cortellis通过众多临床试验及以往多年研究积累,4-1BB 靶点被证实是肿瘤免疫治疗十分有潜力的靶点,目前如何最大化激动作用并最小化毒性是未来4-1BB激动剂研发亟需解决的问题。百奥动物自主研发的4-1BB系列人源化小鼠(详情见下表),是靶向4-1BB抗体开发相关药物进行药效评价的优质模型。B-h4-1BB/h4-1BBL mice高剂量Urelumab对B-h4-1BB小鼠和B-h4-1BB/h4-1BBL小鼠均产生肝毒性用PBS或Urelumab(内部合成)对纯合子B-h4-1BB小鼠和B-h4-1BB/h4-1BBL小鼠进行治疗 (n=5,雌性,7周龄)。第21天采集血清,检测谷丙转氨酶(ALT)、谷草转氨酶(AST)。与PBS对照相比,使用20mg /kg Urelumab治疗B-h4-1BB小鼠的ALT显著升高,而同样的治疗对B-h4-1BB/h4-1BBL小鼠没有影响。与PBS对照相比,B-h4-1BB小鼠AST无明显升高,但20ug/mL urelumab治疗可使B-h4-1BB/h4-1BBL小鼠AST明显升高。在1mg/kg剂量组,B-h4-1BB小鼠和B-h4-1BB/h4-1BBL小鼠的ALT和AST均无明显变化。表明高剂量urelumab对B-h4-1BB和B-h4-1BB/h4-1BBL小鼠有肝毒性作用。数值以平均值±SEM表示。高剂量Urelumab 治疗可导致B-h4-1BB小鼠和B-h4-1BB/h4-1BBL小鼠肝脏淋巴细胞浸润增加肝毒性的病理分析。在B-h4-1BB小鼠(G1-G3)中,当urelumab剂量为1mg/kg时,肝脏未见明显异常变化(G2)。当剂量为20 mg/kg (G3)时,观察到血管周围细胞浸润或肝脏慢性炎症,并伴有轻度病变。在B-h4-1BB/h4-1BBL小鼠(G4-G6)中,1 mg/kg组(G5) 3/5的小鼠出现病理变化(轻度2/5,轻度1/5),而在20 mg/kg剂量(G6)时,所有实验动物肝脏均出现中度变化。总体而言,20 mg/kg组(G6)肝脏病变程度和发生率显著高于1 mg/kg组(G5)。以上结果提示,高剂量(20mg /kg)的urelumab比低剂量(1mg /kg)的urelumab更容易发生血管周围细胞浸润或肝脏慢性炎症。B-h4-1BB/h4-1BBL小鼠对urelumab毒性作用比B-h4-1BB小鼠更敏感。因此B-h4-1BB/h4-1BBL小鼠是较好的临床前毒性评价模型。B-h4-1BB/h4-1BBL药效验证抗人4-1BB抗体在B-h4-1BB/h4-1BBL小鼠中的抗肿瘤药效。(A) 纯合子B-h4-1BB/h4-1BBL小鼠 (雌性,6-8周龄,n=5) 皮下接种小鼠结肠癌MC38细胞系。结果显示抗人4-1BB抗体能够显著抑制MC38肿瘤细胞的生长。(B)治疗期间体重变化。如图A所示,抗人4-1BB抗体在B-h4-1BB/h4-1BBL小鼠中有效地控制肿瘤生长,这表明B-h4-1BB/h4-1BBL小鼠可为体内评价抗人4-1BB抗体药效提供有力的临床前研究模型。值表示为平均值±SEM。参考文献[1].https://mp.weixin.qq.com/s/kvwfybpx63XQn26xdijIZwhttps://mp.weixin.qq.com/s/999cX0RaXHLyXxZ8MhIaBw[2]. Chester C, Sanmamed MF, Wang J, Melero I. Immunotherapy targeting 4-1BB:mechanistic rationale, clinical results, and future strategies. Blood. 2018 Jan4;131(1):49-57. doi: 10.1182/blood-2017-06-741041. Epub 2017 Nov 8. PMID:29118009.

异常的脂质循环和存储是导致心血管疾病和代谢性疾病的风险因素。富含TG(triglycerides,甘油三酯)的脂蛋白(如VLDL,乳糜微粒)在脂蛋白脂肪酶(LPL)的作用下将TG水解成为自由脂肪酸(FFA),进而被不同组织摄取和利用。因此,LPL对于TG代谢和脂肪分布具有重要作用,而LPL的活性受ANGPTL3, 4, 8的调节[1]。ANGPTL(Angiopoietin-like proteins,血管生成素样蛋白)家族包括8个成员,是分泌型糖蛋白,具有3个保守的结构域,即N端的信号肽、coiled-coil结构域(CCD)和C端fibrinogen-like结构域(FLD);只有ANGPTL8例外,它没有C端的FLD。对于ANGPTL3, 4, 8来说,它们都具有LPL和内皮脂肪酶(EL)的抑制结构域(specific epitope1, SE1)。图1. ANGPTL家族蛋白结构域[2]ANGPTL3主要在肝脏中表达,可抑制LPL和EL的酶活性,从而抑制TG水解,也就是抑制VLDL/乳糜微粒向TG含量低的脂蛋白(LDL, HDL)的转换,于是血浆中VLDL水平升高,可引发动脉粥样硬化斑块的产生。另一方面,ANGPTL3的FLD可与α5β3整合素结合,引发斑块新生血管、内膜增厚和炎症等。目前为止,三类ANGPTL3抑制剂药物在开发中,分别是单抗,反义寡核苷酸(ASO)和CRISPR/Cas9基因编辑,它们可促进VLDL的水解。已有的PCSK9抑制剂可促进LDL受体循环,增强肝脏摄取LDL并抑制肝脏分泌VLDL。由于ANGPTL3抑制剂与PCSK9抑制剂的作用机制不同,二者可能具有协同治疗冠心病(CHD)的作用。图2. ANGPTL3的生物学功能及其抑制剂药物的工作机制[3]ANGPTL3的活性依赖ANGPTL8的活化,但相对稳定,不受营养状况的影响。而ANGPTL4和ANGPTL8的水平受进食状况的影响,且呈相反的关系。 禁食或运动状态下,ANGPTL4表达上调(ANGPTL8下调),使WAT中LPL的活性减弱。于是,循环中的TG就进入外周组织(如心脏和肌肉)被利用。 ANGPTL3/8复合物分泌入血,可抑制循环系统、以及氧化组织(如心脏、肌肉)中的LPL活性。喂食状态下,ANGPTL8上调(ANGPTL4下调),使得心脏和肌肉中的LPL活性受ANGPTL3/8抑制,进而使得循环中的TG进入白色脂肪组织(WAT)存储起来。 总的来说,ANGPTLs可调节LPL活性,在禁食条件下促进TG被外周组织的利用,并在进食状态下促进脂质存储[4]。图3. ANGPTL3, 4, 8在不同营养状况下调节血脂的工作机制[1]ANGPTL3,4,8相关药物ANGPTL3是一个值得期待的心血管疾病靶点。2021年2月,FDA批准再生元的依维库单抗(evinacumab, anti-hANGPTL3 antibody)治疗家族性高胆固醇血症,这是此类药物中首个被批准上市的(first-in-class)。依维库单抗可降低甘油三酯,用于治疗纯合子家族性高胆固醇血症(HoFH)患者,增加循环中含apoB的脂蛋白的清除率来降低LDL。另外,还有一些开发靶向ANGPTL3的创新疗法的药物,其中比较领先的是辉瑞从Akcea 和 Ionis处获得全球独家许可的Vupanorsen。Vupanorsen作为ANGPTL3的反义寡核苷酸,能够通过抑制ANGPTL3合成,提高脂蛋白脂肪酶(LPL)的活性,从而降低富含TG脂蛋白(TRL)水平。国内也已开始对ANGPTL靶点进行药物研发,如苏州瑞博生物正在开发靶向ANGPTL3的siRNA抑制剂。活跃在研的ANGPTL8药物只有一款礼来公司的LY-3475766,处于临床I期。这是一款单克隆抗体药物,靶向ANGPTL3/8复合物,可显著降低血脂异常病人血浆中的TG和残存胆固醇含量。靶向ANGPTL4的药物主要有再生元开发的单克隆抗体REGN-1001和Lexicon医药开发的14D12单抗,均用于降血脂。为助力调控脂质代谢的药物开发,百奥动物自主研发了ANGPTL3, 4, 8的人源化小鼠。B-hANGPTL3 mice品系名称:C57BL/6-Angptl3tm1(ANGPTL3)/Bcgen背景:C57BL/6产品编号:112224蛋白表达分析用ELISA分析杂合B-hANGPTL3小鼠中ANGPTL3蛋白的种属特异性表达。取野生型C57BL/6小鼠和杂合B-hANGPTL3小鼠的血浆进行分析。mANGPTL3的表达在两种鼠中均可检测到,而hANGPTL3的表达只在杂合B-hANGPTL3小鼠中检测到,野生型小鼠中检测不到。B-hANGPTL3 mice plus品系名称:C57BL/6-Angptl3tm4(ANGPTL3)/Bcgen背景:C57BL/6产品编号:112523B-hANGPTL3 mice plus小鼠中的mANGPTL3基因(包括3'UTR)被替换为hANGPTL3基因(包括3'UTR),拟用于针对3'UTR设计的核酸药物评估。蛋白表达分析用ELISA分析纯合B-hANGPTL3 mice plus小鼠中ANGPTL3蛋白的种属特异性表达。取野生型C57BL/6小鼠和纯合B-hANGPTL3 mice plus小鼠的血浆进行分析。mANGPTL3的表达在野生型小鼠中可检测到。hANGPTL3的表达只在纯合B-hANGPTL3 mice plus小鼠中检测到,野生型小鼠中检测不到。B-hANGPTL4 mice品系名称:C57BL/6N-Angptl4tm1(ANGPTL4)/Bcgen背景:C57BL/6N产品编号:111085mRNA表达分析用RT-PCR分析纯合B-hANGPTL4小鼠中ANGPTL4基因的种属特异性表达。mANGPTL4 mRNA的表达仅在野生型小鼠的脂肪组织中检测到,而hANGPTL4 mRNA的表达只在纯合B-hANGPTL4小鼠中检测到,野生型小鼠中检测不到。 蛋白表达分析 用western blot分析纯合B-hANGPTL4小鼠中ANGPTL4蛋白的种属特异性表达。取野生型小鼠和纯合B-hANGPTL4小鼠的脂肪组织进行western blot分析。ANGPTL4的表达在两种鼠中均可检测到,因为ANGPTL4可种属交叉识别。并且,对于两种鼠来说,ANGPTL4的表达量在禁食状态下更高。 B-hANGPTL8 mice品系名称:C57BL/6N-Angptl8tm1(ANGPTL8)/Bcgen背景:C57BL/6N产品编号:111230mRNA表达分析 用RT-PCR分析纯合B-hANGPTL8小鼠中ANGPTL8基因的种属特异性表达。mANGPTL8 mRNA的表达仅在野生型小鼠中检测到,而hANGPTL8 mRNA的表达只在纯合B-hANGPTL8小鼠中检测到,野生型小鼠中检测不到。蛋白表达分析 用ELISA分析纯合B-hANGPTL8小鼠中ANGPTL8蛋白的种属特异性表达。取野生型C57BL/6小鼠和纯合B-hANGPTL8小鼠的血浆进行ELISA分析。ANGPTL8的表达在两种鼠中均可检测到,因为抗体可种属交叉识别。参考文献1. Sylvers-Davie KL, Davies BSJ. Regulation of lipoprotein metabolism by ANGPTL3, ANGPTL4, and ANGPTL8. Am J Physiol Endocrinol Metab. 2021;321(4):E493-E508. doi:10.1152/ajpendo.00195.20212. Carbone C, Piro G, Merz V, et al. Angiopoietin-Like Proteins in Angiogenesis, Inflammation and Cancer. Int J Mol Sci. 2018;19(2):431. Published 2018 Feb 1. doi:10.3390/ijms190204313. Rhee JW, Wu JC. Dyslipidaemia: In vivo genome editing of ANGPTL3: a therapy for atherosclerosis?. Nat Rev Cardiol. 2018;15(5):259-260. doi:10.1038/nrcardio.2018.384. Aryal, Binod et al. “ANGPTL4 in Metabolic and Cardiovascular Disease.” Trends in molecular medicine vol. 25,8 (2019): 723-734. doi:10.1016/j.molmed.2019.05.010



